Riassunto
Viene descritta la composizione floristica e la vegetazione di un’area nel comune di Mogliano Veneto (TV), predisposta dal Consorzio Acque Risorgive ai fini di contenere le acque di piena del rio Rusteghin e di operare la fitodepurazione.
Lo studio si è svolto tra la primavera del 2016 e l’autunno del 2017 e ha dato la possibilità di osservare l’insediamento delle prime specie pioniere su un terreno privo di copertura vegetale fino alla situazione attuale con 137 specie e una discreta copertura da parte delle fitocenosi con caratteristiche idro-igrofile.
Abstract
Floristic analisys and research about the development and evolution of hygrophilous vegetation in a new–formed wetland in Campocroce (Mogliano Veneto – TV)
The floristic composition and the aquatic vegetation of a wetland in the Mogliano Veneto (TV) municipality are described. The area was digged by Consorzio Acque Risorgive to obtain 2 basins in order to contain the Rusteghin ditch flood waters and to carry out a green constructed wetland. The research, carried out from spring 2016 to autumn 2017, gave the chance to study the development of hygrophilous vegetation and the increase in the number of species (from 0 till 137).
Introduzione
Il lavoro botanico oggetto di questo articolo nasce dalla volontà di studiare dal punto di vista floristico e dinamico vegetale la neo formata zona umida in località Campocroce di Mogliano Veneto. Il lavoro di escavazione e di movimento terra per la realizzazione di una zona di fitodepurazione, da parte dell’ente di Bonifica Acque Risorgive, ha infatti determinato un punto zero per quanto riguarda la copertura delle specie vegetali con la presenza di terreni fangosi scoperti.
In questo caso lo studio delle tipologie botaniche delle zone più vicine all’acqua ha come obiettivo secondario quello di definire la possibilità o meno di effettuare gli impianti di specie utili alla fitodepurazione, previsti normalmente dai progetti di questo tipo (Blasi & Paolella, 1992; Borin & Tocchetto, 2007; Gerdovic, 1992; Moroni et al., 1996; Vismara, 1998; Vymazal, 2011, 2013) e che, in accordo con la proprietà, non sono state realizzati proprio per permettere l’analisi floristica.
Inoltre questa indagine potrà contribuire in un’ottica multidisciplinare a definire quali siano le migliori condizioni per il realizzarsi di un assetto vegetazionale completo e utile, decidendo, oltre all’uso o meno di piantumazioni artificiali, il tipo di suolo per le diverse coperture, le altezze delle diverse superfici o la profondità dello specchio d’acqua.
Area di studio
L’area presa in esame si trova nella località di Campocroce nel comune di Mogliano Veneto. È compresa tra una serie di anse del fiume Zero a sud, il Rio Rusteghin ad ovest e la linea ferroviaria Venezia–Treviso ad est. Occupa la parte centrale di due rotatorie della provinciale 65 per Zero Branco. Ha un’area di circa 37.600m2 e le sue coordinate sono longitudine (Roma, M. Mario) 12°3’45”, latitudine 45°29’33” (Fig. 1a).
collocazione del sito
nel comune di Mogliano
Immagine satellitare anno 2005 
immagine satellitare anno 2016
Il terreno prima del progetto di trasformazione dell’area era interessato dalla presenza di un’area agricola coltivata in modo intensivo con mais e soia. A seguito della realizzazione dei lavori di scavo è stata realizzata anche una seconda rotatoria (Fig. 1b-c).
L’impianto di fitodepurazione di Campocroce è stato realizzato secondo il modello a scorrimento superficiale (free water system, FWS) (Veneto Agricoltura, 2014; Ispra, 2012), con lagunaggi a pelo libero d’acqua, nei quali le piante possono radicare in un terreno limoso-argilloso.
Sono state realizzate due vasche di contenimento (bacino A ad ovest, bacino B ad est) e si è costituita un’area a vari livelli altimetrici rispetto al piano campagna circostante (circa 8 m sul livello del mare): un piano elevato a 7,5 m s.l.m., una zona intermedia per la vegetazione delle elofite (golene) a circa 6,5 m s.l.m., e infine i bacini con un fondale a 6 m s.l.m., dunque con una profondità massima di circa 50 cm.
Per rallentare il flusso dell’acqua sono state progettate due penisole che tagliano trasversalmente gli specchi d’acqua, utilizzando due burghe poi ricoperte di terreno. Queste strutture hanno anche il merito di creare un effetto più naturale, similmente ad un fiume con i suoi meandri.
I bacini impermeabilizzati hanno un livello d’acqua costantemente mantenuto da chiuse regolabili. La regolazione del livello idrico è fondamentale nel guidare la formazione dell’habitat in riferimento agli obiettivi ambientali-funzionali.
L’analisi botanica si è concentrata esclusivamente sulle zone sommerse e sui pantani a pelo d’acqua, costituiti appositamente per ospitare la componente floristica legata agli ambienti acquatici e semiacquatici.
Esse vengono qui denominate “golene” per l’analogia morfologica che hanno con le banchine alluvionali che si formano in maniera naturale nei corsi d’acqua in genere. Sono in parte sommerse per qualche centimetro (Fig. 2, in azzurro) e in parte tenute appena al di sopra del livello acqueo (Fig. 2 in verde).
Il terreno risulta quindi generalmente ben bagnato durante tutto l’anno. Tutte le golene sono costituite da materiale limoso-argilloso. Nel caso di forti piogge possono formarsi delle pozze temporanee di qualche centimetro di profondità in alcune bassure o provocare l’innalzamento per qualche giorno dell’intero bacino, sommergendo tutta la superficie. All’opposto nel pieno dell’estate nelle golene più ampie si possono costituire delle zone di terreno asciutto. Le condizioni del substrato si riflettono sulla componente floristica, evidenziando brevi oscillazioni nella presenza di specie igrofile a specie xeriche.
Nella rotonda di Campocroce queste aree si sviluppano sia nel bacino A sia nel bacino B. per un totale di circa 13.900 m2.
Le golene nel bacino A coprono circa 2050 m2. La parte sommersa corrisponde a circa 450 m2 e durante i due anni di studi è rimasta per la maggior parte priva di vegetazione. L’area è stata suddivisa in quattro zone che corrispondono alle prime quattro aree di rilevamento floristico (Fig. 2a). Le golene del bacino B coprono circa 11.850 m2. La parte sommersa ha una superficie di 2322 m2, pari a circa il 20% del totale.
Nel secondo bacino, più esteso e diversificato, sono state definite altre 14 aree di rilevamento (Fig. 2b)
La flora
La campagna di raccolta si è svolta tra la primavera del 2016 e l’autunno del 2017, in modo da osservare la fenologia di tutte le specie vegetali presenti nel sito e l’evoluzione in termini di comparsa/scomparsa e abbondanza nei due anni. In particolare sono stati eseguiti un totale di 29 sopralluoghi: 14 nel 2016 (tra il 28 Aprile e il 26 Ottobre), 15 nel 2017 (tra il 13 Aprile e il 14 Settembre). Nei due anni di ricerca si sono così individuati tre periodi di osservazione (primaverile, estivo ed autunnale), che indicheremo per semplificazione come: Primo Periodo (pp), Secondo Periodo (sp), Terzo Periodo (tp), in 18 zone abbastanza ben delimitate in cui eseguire i rilevamenti floristici (Figg. 2a, 2b; Tab 1).
Un rilievo viene identificato dall’anno, dal periodo in cui è stato eseguito e dal numero (es. 2016 pp ril.1 corrisponde al primo rilievo in assoluto). Nei rilievi effettuati si è tenuto conto sia della presenza che della copertura delle specie (Pignatti, 1994, 1998).
Sono state raccolte le indicazioni sulla forma biologica delle specie presenti (Pignatti, 1982, Raunkiaer, 1934), rapportandola alla scala temporale per individuare uno schema nella successione floristica, e si è ritenuto utile associare ad ogni entità vegetale i propri indici ecologici (Ellenberg, 1988; Ellenberg et al., 1992).

a) localizzazione dei rilievi
nel bacino A – numeri 1-4;
b) localizzazione dei rilievi nel bacino B – numeri 5-18;
fondo verde – zona emersa; fondo azzurro – zona sommersa.
L’elenco floristico che segue conta 137 specie, incluse in 93 generi e 36 famiglie, determinate secondo la “Flora d’Italia” (Pignatti, 1982), con l’ausilio di altri testi come “Flora Italica” (Zangheri, 1976).
Equisetaceae: Equisetum hyemale L., Equisetum palustre L., Equisetum arvense L., Equisetum telmateja Ehrh..
Salicaceae: Salix alba L., Salix cinerea L., Salix purpurea L., Populus alba L., Populus nigra L..
Urticaceae: Parietaria officinalis L..
Polygonaceae: Polygonum aviculare L., Polygonum mite Schrank, Polygonum lapathifolium L., Polygonum persicaria L., Rumex acetosa L., Rumex crispus L..
Amaranthaceae: Amaranthus deflexus L..
Portulacaceae: Portulaca oleracea L..
Caryophyllaceae: Stellaria neglecta Weihe, Cerastium glomeratum Thuill., Myosoton aquaticum (L.) Moench, Velezia rigida L..
Ranunculaceae: Ranunculus acris L., Ranunculus repens L., Ranunculus bulbosus L., Ranunculus sceleratus L..
Guttiferae: Hypericum perforatum L..
Cruciferae: Arabidopsis thaliana (L.) Heynh., Rorippa sylvestris (L.) Besser, Capsella bursa-pastoris (L.)Moe., Brassica napus L..
Platanaceae: Platanus hybrida Brot..
Rosaceae: Potentilla reptans L..
Leguminosae: Medicago lupulina L., Trifolium repens L., Trifolium pratense L., Lotus corniculatus L..
Oxalidaceae: Oxalis fontana Bunge.
Euphorbiaceae: Euphorbia helioscopia L.
Malvaceae: Abutilon theophrasti Medicus.
Tamaricaceae: Tamarix gallica L..
Lythraceae: Lythrum salicaria L..
Onagraceae: Oenothera biennis L., Epilobium hirsutum L., Epilobium parviflorum Schreber, Epilobium tetragonum L..
Umbelliferae: Berula erecta (Hudson) Coville, Daucus carota L..
Primulaceae: Lysimachia nummularia L., Lysimachia vulgaris L., Anagallis arvensis L., Samolus valerandi L..
Gentianaceae: Blackstonia perfoliata (L.)Hud., Centaurium pulchellum(Swartz) Druce.
Rubiaceae: Galium album Miller, Galium aparine L..
Convolvulaceae: Cuscuta campestris Yuncker, Convolvulus arvensis L..
Boraginaceae: Symphytum officinale L., Myosotis scorpioides L..
Labiatae: Scutellaria galericulata L., Stachys palustris L., Prunella vulgaris L., Lycopus europaeus L., Mentha aquatica L..
Scrophulariaceae: Gratiola officinalis L., Veronica peregrina L., Veronica anagallis aquatica L..
Plantaginaceae: Plantago major L., Plantago lanceolata L..
Compositae: Eupatorium cannabinum L., Aster novi-belgii L., Aster lanceolatus Willd., Aster squamatus (Sprengel) Hieron., Conyza bonariensis (L.) Cronq., Conyza canadensis (L.)Cronq., Erigeron annuus (L.)Pers., Bidens tripartita L., Ambrosia artemisiifolia L., Matricaria chamomilla L., Leucanthemum vulgare Lam., Artemisia vulgaris L., Tussilago farfara L., Senecio vulgaris L., Arctium lappa L., Cirsium vulgare (Savi)Ten., Cirsium arvense (L.)Scop., Centaurea nigrescens Wild., Picris hieracioides L., Picris echioides L., Taraxacum officinale Weber, Sonchus asper (L.)Hill, .
Alismataceae: Alisma plantago-aquatica L..
Juncaceae: Juncus bufonius L., Juncus compressus Jacq, Juncus gerardi Loisel., Juncus effusus L., Juncus inflexus L..
Graminaceae: Poa annua L., Poa trivialis L., Festuca pratensis Hudson, Festuca arundinacea Schreber, Glyceria maxima (Hartman)Holberg, Lolium multiflorum Lam., Bromus hordeaceus L., Agropyron repens (L.)Beauv., Phragmites australis (Cav.) Trin, Typhoides arundinacea L.(Moench), Alopecurus myosuroides 36
Hud., Cynodon dactylon (L.)Pers., Panicum capillare L., Panicum dichotomiflorum Michx, Echinochloa crus-galli (L.)Beauv, Paspalum dilatatum Poiret., Paspalum paspaloides (Michx)Scrib., Setaria viridis (L.)Beauv., Sorghum halepense (L)Pers.
Lemnaceae: Lemna minor L..
Sparganiaceae: Sparganium erectum L..
Typhaceae: Typha latifolia L., Typha laxmanii Lepechin.
Cyperaceae: Carex contigua Hoppe, Carex otrubae Podp., Carex elata All., Carex panicea L., Carex distans L., Carex pseudocyperus L., Carex acutiformis Ehrh., Carex hirta L., Bolboschoenus maritimus (L.)Palla, Schoenoplectus lacustris (L.) Palla, Cyperus longus L., Cyperus esculentus L., Cyperus glomeratus L., Cyperus fuscus L., Cyperus flavescens L..
L’elenco rispecchia la situazione floristica complessiva alla fine della seconda stagione di rilevamento (autunno 2017). Si nota l’alto numero di specie della famiglia Cyperaceae fortemente legata agli ambienti acquatici e semiacquatici (15 specie), con i generi Carex e Cyperus, e della famiglia Juncaceae con il genere Juncus (5 specie).
Analizzando i diversi ambienti di crescita (Pignatti, 1982) (Fig. 3) si evidenzia una divisione tra le specie di ambiente umido (dal generico “ambienti umidi” a “fossi e stagni” ad “acqua dolce”) e le specie sinantropiche legate alle aree molto disturbate, ruderali e degli incolti. Questa situazione floristica rispecchia appieno la condizione attuale di questa zona: da una parte un neoambiente adatto allo sviluppo delle specie igrofile, dall’altra un ecosistema ancora immaturo e influenzato da specie dal rapido sviluppo capaci di approfittare del terreno nudo anche in situazioni più aride, come può avvenire in estate, o, come è avvenuto, nel primo periodo del primo anno.

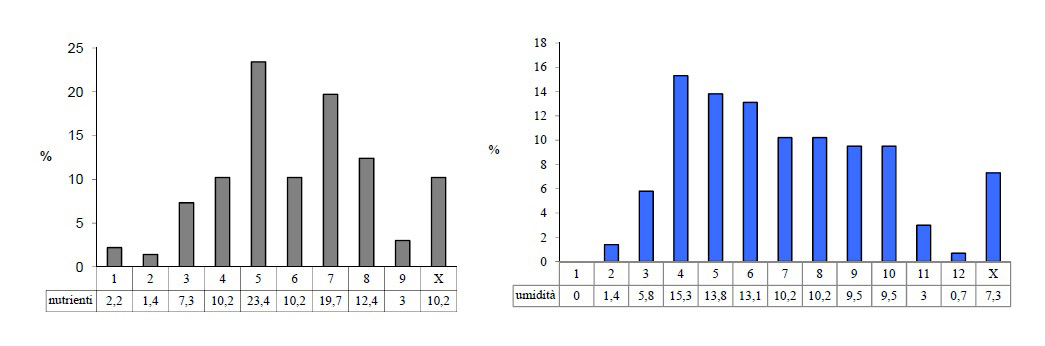
Questo aspetto è comprovato anche dall’osservazione dei valori di bioindicazione, che riguardano il fattore “umidità” e “nutrienti” (Fig.4). Risulta una flora con una percentuale elevata di specie tipiche di ambiente umido, circa il 63% e che per il 70% è adattata a vivere in un ambiente da mediamente ricco a molto ricco di nutrienti, rispecchiando quindi quella che è la volontà progettuale di realizzazione di quest’area, cioè essenzialmente un ambiente acquatico di accumulo e trasformazione di nitrati e fosfati.
Confrontando l’elenco floristico con il manuale dell’ISPRA (2012) riguardo alla “Progettazione e realizzazione di impianti di fitodepurazione per il trattamento delle acque reflue urbane” e con la pubblicazione di Veneto Agricoltura (2014) “La fitodepurazione per il trattamento di acque di origine agricola e di reflui zootecnici” , si evidenzia che nelle tabelle che indicano le specie vegetali utilizzate negli impianti FWS in Italia sono elencate 14 specie di elofite e 1 specie pleustofitica presenti anche a Campocroce. Insieme a queste specie di elevato valore morfologico-funzionale ne sono presenti altre altrettanto significative da un punto di vista ecologico più ampio.
Typha latifolia: trova nelle golene di Campocroce un habitat ideale. Infatti è la pianta più diffusa in zona comparendo in quasi tutti i rilievi già dal primo periodo del primo anno, prima nel bacino B e poi nel bacino A, con coperture elevate.
Typha laxmannii: è considerata specie rara (Pignatti, 1982) e indicata nella Lista rossa regionale delle piante vascolari del Veneto (Buffa et al, 2016) come EN per la regione. Accompagna e si mischia a T. latifolia fin dal primo anno. Nel terzo periodo del secondo anno ha rafforzato la sua presenza, formando densi popolamenti in quattro diversi rilievi (Fig. 5).
Phragmites australis: nell’area esaminata non ha ancora una presenza decisiva ed è stata rilevata solo nel secondo anno prima nel bacino B poi nel bacino A, sempre con coperture inferiori all’ 1%. Sembra quindi che non si siano ancora realizzate le condizioni ecologiche migliori per il suo sviluppo, probabilmente anche per la maggiore adattabilità ai suoli nudi delle specie dominanti.

a – visione di insieme 
b – particolare dell’infiorescenza 
c -particolare dei frutti (nucule)

Schoenoplectus lacustris: è stata rilevata dal secondo periodo del secondo anno in due rilievi e dal terzo periodo si è diffusa in un’altra zona, ma sempre con coperture ridotte, tra 1 e 5 %.
Juncus spp: nella zona studiata sono state trovate 5 specie. Juncus bufonius è stata rilevato in numerosi rilievi fin dal primo periodo del primo anno soprattutto nelle golene del bacino B. Juncus effusus e Juncus inflexus sono piante più grandi e sono state trovate esemplari isolati a partire dal secondo anno solo nel bacino B. Juncus gerardi è una presenza atipica (infatti è dato assente per la provincia di Treviso nella lista rossa regionale delle piante vascolari del Veneto) (Buffa et al. 2016). Sono stati trovati alcuni esemplari nelle golene del bacino B (appendice Aree di rilevamento 10, 11, 9) dal terzo periodo del secondo anno. Juncus compressus è la specie che ha avuto più successo. Presente fin dal primo anno si è diffusa velocemente in tutta l’area fino a raggiungere coperture notevoli: tra il 20 e 40% in più rilievi.
Carex spp.: sono state rilevate otto specie appartenenti al genere Carex, tutte a partire dal secondo anno di rilevamenti. Tra queste Carex elata si è diffusa velocemente, raggiungendo tra le Carex la maggiore copertura. All’opposto C. hirta, trovata con pochi esemplari in un unico rilievo nel bacino B. Le altre specie in ordine di diffusione sono C. otrubae, C. panicea, C. acutiformis, C. pseudocyperus (segnalata nella lista rossa regionale delle piante vascolari del Veneto come specie CR) (Buffa et al. 2016), C. contigua, C. distans.
Sparganium erectum: specie ancora poco diffusa. Raggiunge la copertura maggiore solo nel rilievo 2, mentre nel resto dell’area ha una presenza frammentata.
Bolboschoenus maritimus: tende a formare popolamenti puri e infatti si trova sempre aggregata in gruppi numerosi monospecifici. Predilige acque poco profonde e zone assolate. Lo troviamo sia nel bacino A che B già dal primo anno di rilevamenti. Si è poi diffuso notevolmente nel secondo anno raggiungendo buone coperture.
Cyperus spp: il genere Cyperus ha trovato a Campocroce ampia diffusione. Le 5 specie rilevate sono un numero elevato se rapportato all’estensione dell’area. Soprattutto Cyperus fuscus che a partire dal secondo periodo del primo anno è presente in molti rilievi di entrambi i bacini, seguita da Cyperus flavescens e Cyperus glomeratus. Nel secondo anno questa tendenza è stata confermata e la specie ha raggiunto notevoli coperture. Nel primo anno si è riscontrato anche Cyperus longus, rimasto relegato nel rilievo 2 e, alla fine del secondo anno, Cyperus esculentus. Probabilmente la mancanza di una copertura totale da parte delle piante più alte ha permesso la diffusione delle piccole specie di Cyperus, lasciando al suolo una buona areazione e disponibilità di luce.
Lemna minor: è stata trovata dal primo periodo del secondo anno prima nel bacino A e poi nel B, ma con minor successo, a ridosso delle rive tra la tifa e lo sparganio, protetta dal flusso dell’acqua. Infatti è sufficiente una debole corrente per trasportare le piantine altrove e finché le golene sommerse non saranno coperte dalla vegetazione non avrà lo sviluppo desiderato. È l’unica idrofita natante. Non è da escludere la possibilità in futuro di immettere nei bacini attraverso la struttura di inlet acqua dallo Zero e con essa probabili piante acquatiche non presenti nel fosso Rusteghin. Infatti sono state rinvenute nei pressi della chiusa nella parte di collegamento con lo Zero (fuori area di rilevamento) due specie che potrebbero dare un forte contributo al processo di fitodepurazione: Ceratophyllum demersum L. ed Elodea canadensis Michx.
La vegetazione
Dal punto di vista vegetazionale le golene vengono qui analizzate come un ambiente essenzialmente uniforme. Questa uniformità è sintomo della giovane età, oltre che della condivisione della stessa origine e dell’influenza dei medesimi fattori biotici. Tuttavia si scorgono delle probabili tendenze evolutive, dovute a leggere differenze: altimetria, vicinanza o lontananza dalla strada, sponde o altri canali preesistenti, esposizione.
I rilievi floristici effettuati nell’ultimo anno di analisi dell’area, al termine del periodo di campionamento (estate 2017), sono stati tabulati per tipologie affini in modo da poter realizzare uno studio delle possibili associazioni vegetali che si sono formate. Tali fitocenosi sono state inquadrate secondo l’impostazione della fitosociologia classica (Associazioni, Alleanze, Ordini e Classi) (Braun Blanquet, 1928), tenendo conto che il tempo di sviluppo è stato limitato e quindi si tratta ancora di cenosi pioniere e non ancora del tutto consolidate e che si presentano in strutture a mosaico, a seconda dell’influenza dei diversi fattori ecologici. 40
Ciò che risulta è un quadro che mostra una vegetazione già con una discreta copertura e una buona presenza di diverse specie elofitiche. Al momento la struttura della vegetazione, in fase di completamento e di organizzazione e di raggiungimento di un equilibrio, è stata presentata solo a livello di syntaxa ad ampio spettro senza poter scendere al livello di associazione vegetale, in quanto le specie registrate sono per lo più attribuibili a livelli sintassonomici superiori. L’inquadramento sintassonomico adottato fa riferimento ai lavori di vari autori: Oberdorfer (1977, 1978, 1983, 1992) e Pignatti (1998) ed inoltre, per una revisione finale a http://www.prodromo-vegetazione-italia.org/. Per completare l’analisi vegetazionale presentiamo il quadro sintassonomico, la tabella 1 che indica la presenza/copertura delle specie di interesse fitosociologico del secondo periodo del secondo anno e alcune osservazioni riguardanti le diverse classi vegetazionali.
Quadro sintassonomico
Classe Bidentetea tripartitae Tx., Lohm. Et Prsg ex von Rochow 51
Ordine Bidentetalia tripartitae Br.-Bl. & Tuxen ex Klika in Klika & Hadac 1944
Alleanza Bidention tripartitae Nordh. 40
Classe Isoeto-Nanojuncetea Br.-Bl.et Tx. Ex Westhoff, Dijk & Passchier 1946
Ordine Nanocyperetalia flavescentis Klika 1935
Alleanza Nanocyperion flavescentis W. Koch ex Libbert 1932
Classe Phragmito australis-Magnocaricetea elatae Klika in Klika et Novak 41
Ordine Phragmitetalia australis W. Koch 26
Alleanza Phragmition communis W. Koch 26
Ordine Magnocaricetalia elatae Pignatti 1953
Alleanza Magnocaricion elatae W. Koch 26
Ordine Nasturtio officinalis-Glycerietalia fluitantis Pignatti 1953
Alleanza Glycerio fluitantis-Sparganion neglecti Br.-Bl. et Siss in Boer 42
Classe Salicetea purpureae Moor 58
Ordine Salicetalia purpureae Moor 58
Alleanza Salicion albae Soó 30
Classe Querco roboris-Fagetea sylvaticae Br.-Bl. et Vlieg. In Vlieg. 37
Ordine Fagetalia sylvaticae Pawl. 28 in Pawl., Sok. Et Wall. 28
Alleanza Alnion incanae Pawlowski in Pawloski & Wallisch 1928
Classe Bidentetea tripartitae Tx., Lohm. Et Prsg ex von Rochow 51
In questi ambienti caratterizzati da acque scure, ricche di sostanze nutritive con presenza di nitrati e nei terreni impregnati, si sviluppano diverse specie, appartenenti a questa classe (specie tipicamente nitrofile) ed in particolare si segnalano le specie caratteristiche dell’alleanza Bidention tripartitae.
Classe Isoeto-Nanojuncetea Br.-Bl.et Tx. Ex Westhoff, Dijk & Passchier 1946
Questa classe è rappresentata a Campocroce con l’alleanza Nanocyperion. In generale è caratterizzata da specie più “basse”, e sovente con una maggiore necessità di umidità nel terreno, più prossime all’acqua o comunque in bassure o depressioni all’interno delle golene lungo i bacini.
Classe Phragmito australis-Magnocaricetea elatae Klika in Klika et Novak 41
A questa classe fitosociologica va ascritto il maggior numero di specie. Essa raggruppa le tipiche associazioni ad elofite (i canneti anfibi) che costeggiano i corsi d’acqua a debole pendenza e le bordure di stagni, laghi e ambienti umidi.
Tab. 1. Tabella di presenza/copertura delle specie di interesse fitosociologico (secondo periodo del secondo anno).
All’interno di tale sintaxon è stato possibile evidenziare un contingente di specie che possono essere attribuite al livello di tre diverse e specifiche alleanze ed in particolare l’alleanza Phragmition australis, l’alleanza Caricion elatae, ed infine l’alleanza Sparganio-Glycerion fluitantis.
Classi Salicetea purpureae Moor 58 e Querco-Fagetea Br.-Bl. et Vlieg. In Vlieg. 37
Nel corso dei due anni di studio è stato possibile osservare la colonizzazione massiccia delle aree meno soggette ad inondazione da parte di specie arbustive ed arboree, inizialmente in forma di plantule ma, al termine del secondo anno, anche in forma di cespugli già ben sviluppati, soprattutto di quelle specie che presentano un ritmo di crescita piuttosto vigoroso (salici). Tali specie vengono ad essere aggruppate nelle classi fitosociologiche Querco-Fagetea e Salicetea purpureae con l’alleanza Salicion albae. I contingenti di specie arbustive ed arboree danno anche l’idea di come potrebbe evolvere nel corso degli anni l’ambiente ripariale, soprattutto in aree più distali rispetto al pelo dell’acqua.
A queste specie che abbiamo indicato come caratteristiche di alcuni aggruppamenti specifici e ben identificabili, in quanto complessivamente ben rappresentati sia come numero di specie che come coperture percentuali del terreno, sono da registrare altre specie che presentano una maggiore ampiezza di condizioni ecologiche e maggiore tolleranza alle variazioni del substrato. Tuttavia anche queste specie si adattano a vivere a condizioni di zone golenali e in prossimità delle sponde dei corsi d’acqua. Nel presente lavoro sono state indicate come specie compagne:
Tra le specie compagne è da segnalare la presenza dell’unica specie che si caratterizza per una forma biologica completamente diversa dalle altre fin qui analizzate, cioè Lemna minor che appartiene alla classe fitosociologica Lemnetea minoris. In effetti è evidente la mancanza di specie tipicamente acquatiche sommerse, radicanti al fondo o galleggianti se si esclude la presenza di più o meno fitte matasse di alghe unicellulari filamentose del genere Spirogyra (ordine Zygnematales) che spesso accompagna i corpi idrici a forte eutrofizzazione.
Dinamica vegetale
Una delle finalità della ricerca effettuata a Campocroce è capire dal punto di vista floristico quale sarà il futuro di questo ambiente. Possono essere utili alcune osservazioni preliminari. Rapportando la percentuale media di copertura nei 6 periodi totali fra il 2016 e il 2017 con la relativa media del numero di specie, si nota che gli andamenti delle due linee tendono a divergere (Fig. 6).
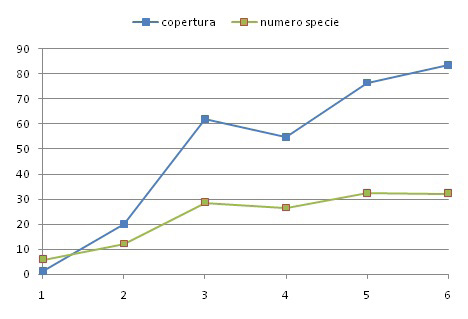
Le curve hanno la pendenza maggiore nel periodo estate-autunno 2016, in cui vi è stata la veloce colonizzazione di un terreno praticamente ancora nudo; vi è una flessione negativa nel periodo invernale, dovuta alla scomparsa delle specie annuali (terofite), a cui segue un ulteriore incremento ma con una pendenza minore – nell’estate-autunno 2017 –. Questo può indicare l’avvicinarsi della “potenzialità floristica” di questo ambiente, che si manifesta con un calo nell’aumento del numero di specie, seguito da un ampliamento delle entità più adattate, fino al raggiungimento del 100% di copertura. Se si mantengono le condizioni ambientali costanti, è prevedibile, nel prossimo futuro, un assestamento della struttura floristica con l’arrivo di poche specie dovuto per lo più ai fenomeni di comparsa-scomparsa, alla stagionalità o all’arrivo di specie più specializzate o di invasive alloctone, fino al raggiungimento di una struttura vegetazionale stabile probabilmente riconoscibile a livello di associazione fitosociologia. È molto probabile che lo sviluppo futuro di alcune specie che tendono a formare consorzi monospecifici diminuirà la biodiversità generale dell’area, come ad esempio tende a fare Phragmites australis.
Confrontando gli spettri biologici nei tre periodi nei due anni di studio si possono fare altre osservazioni (Fig. 7).

biologici nel tempo:
pp = primo periodo (primavera)
sp = secondo periodo (estate)
tp = terzo periodo (autunno).
Per ogni singola forma biologica:
T = terofite
H = emicriptofite
G = geofite
I = igrofite
P = fanerofite
Ch = camefite
He = elofite.
Le terofite (T) sono rappresentate per la maggior parte da specie erbacee annuali pioniere, tipiche anche di ambienti ruderali disturbati. Il fatto di passare la stagione avversa sotto forma di seme permette a queste piante di essere tra le prime a colonizzare nuovi territori, soprattutto se, come nel nostro caso, creati ex novo in seguito a spostamento e risistemazione del terreno. La loro percentuale nel corso dei due anni è costantemente in calo, conseguenza dello sviluppo delle piante perenni.
Quella delle emicriptofite (H) solitamente è la forma biologica tipica della flora delle nostre latitudini. La sua percentuale costante nel tempo indica l’appartenenza della zona di studio alla fascia temperata dell’Italia continentale
Alle geofite (G) appartengono le specie che abbiamo riscontrato di maggior successo (Typha latifolia, Typha laxmannii, Bolboschoenus maritimus, Juncus compressus), che presentano gemme sotto il terreno in organi sotterranei come bulbi, tuberi e rizomi. Molto simili sono le piante elofite (He) in senso stretto. Anch’esse portano le gemme sotto il terreno in radici ingrossate sommerse dall’acqua, però mantengono gli organi aerei (fusto e foglie) durante l’inverno. Nel nostro caso si sono sviluppate alla fine del secondo anno (Schoenoplectus lacustris, Cyperus glomeratus, Cyperus esculentus).
Le fanerofite (P) sono piante legnose, arboree o arbustive, che a Campocroce sono rappresentate da specie adattate agli ambienti umidi (Salix alba, Salix cinerea, Salix purpurea, Populus alba, Populus nigra, Tamarix gallica). I primi esemplari sono arrivati alla fine del primo anno e hanno confermato la loro presenza nel secondo.
Le idrofite (I) sono solitamente le specie meno diffuse, ma vista la tipologia di ambiente indagato, potrebbe essere la forma biologica che nel tempo subirà l’aumento maggiore. Tra queste ricordiamo in particolare Lemna minor, che è specie già presente nel sito.
In considerazione di quanto affermato precedentemente è probabile che lo spettro biologico del terzo periodo del 2017, rappresenti a grandi linee la situazione che si manterrà nei prossimi anni.
Considerando questi dati e con l’aiuto di immagini che testimoniano le modificazioni dell’aspetto floristico delle diverse zone, è possibile descrivere l’evoluzione floristica dell’intera area. Analizziamo una di queste zone.
Rilievo/Zona 1
Il rilievo 1 o zona 1 ha un’estensione di 624 m2. Non presenta porzioni volutamente sommerse, ma alcune pozze di pochi cm. Nel primo periodo (Aprile 2016) conta 10 specie, alcune tipicamente prative (Alopecurus myosuroides, Capsella bursa pastoris), altre di ambiente umido (Glyceria maxima, Cyperus fuscus, Veronica anagallis aquatica) con copertura dell’1%. Alla fine di Giugno l’area si arricchisce di specie, mantenendo la dicotomia tra quelle pioniere (Poa annua, Plantago major) e quelle idrofile (Ranunculus sceleratus, Alisma plantago aquatica). La prima stagione si chiude con un alto numero di specie (32) e una copertura notevole (40%), tra cui però spiccano le specie ruderali Echinocloa crus-galli, Panicum dicotomiflorum e Bidens tripartita.
A primavera del 2017 la zona si presenta ricoperta del materiale vegetale delle specie annuali dell’anno precedente e la prima sorpresa viene dalla vegetazione delle pozze con l’affermazione del Bolboschenus maritimus, che da qui si diffonderà nel bacino A e poi nel B. Rafforzano la loro copertura anche altre specie importanti (Typha latifolia), altre compaiono (Typha laxmannii, Berula erecta, Juncus compressus), arrivano le prime fanerofite (Salix purpurea, Salix cinerea) e alcune specie prative reptanti corrono sul terreno lasciato spoglio (Trifolium repens, Ranunculus repens).
Nell’estate del 2017 non si assiste più alla veloce diffusione delle specie ruderali, ma alla rapida diffusione della tifa e delle altre specie idrofile, arrivate in primavera o nell’anno 2016 (Lycopus europaeus, Mentha aquatica, Cyperus fuscus, Veronica anagallis aquatica). Permangono specie di prato ma con coperture basse.
La fine della stagione di rilevamenti descrive questa zona come un ambiente umido con una componente floristica spiccatamente idrofila, in cui le specie nitrofile, ruderali e terofite prative prima dominanti o sono scomparse o sono diminuite drasticamente in copertura. Alla fine si contano 49 specie con una copertura del 90 % (Fig. 8).

A) pp 2016 = primo periodo (primavera) 2016 
B) sp 2016 = secondo periodo (estate) 2016 
C) tp 2016 = terzo periodo (autunno) 2016 
D) pp 2017 = primo periodo (primavera) 2017 
E) sp 2017 = secondo periodo (estate) 2017 
F) tp 2017 = terzo periodo (autunno) 2017
Conclusioni
L’analisi floristica e vegetazionale della rotonda di Campocroce ha messo in evidenza le potenzialità di questo ambiente. Pur essendo un’area ridotta e di recente formazione ha dimostrato di possedere le caratteristiche adeguate allo sviluppo di un manto vegetale ricco con una notevole biodiversità.
Il numero di specie complessive ha raggiunto un considerevole valore in rapporto alla superficie indagata: 137 specie per 13900 m2. Anche la tipologia delle specie presenti è mutata nel corso del biennio, in quanto se nel primo anno abbondavano le specie prative, infestanti e ruderali, nel secondo si sono sviluppate le specie più tipiche dell’habitat acquatico. Questa evoluzione si può attribuire in parte al completamento dei lavori idraulici alla fine del primo anno, che ha consentito nel secondo un livello dell’acqua leggermente superiore, in parte al taglio delle erbe del piano campagna e alle opere di movimento terra (per l’impianto di alberi e arbusti), che hanno ridotto notevolmente l’apporto di semi delle specie prative annuali terofite, in parte alla stessa evoluzione delle piante perenni idrofite, che richiede dei tempi più lunghi di attecchimento e di diffusione.
A questo proposito sarebbe importante continuare la ricerca sulla dinamica floristica e vegetazionale nei prossimi anni, perché è probabile che si sviluppino quelle specie che ancora non sono state osservate ed in particolare le idrofite natanti e radicanti. La comparsa di queste e altre specie, per il momento assenti, potrebbe anche portare alla “chiusura” dei consorzi vegetali andando a costituire delle vere e proprie associazioni fitosociologiche.
Infatti una situazione simile è descritta in lavori svolti in passato in ambienti affini per tipologia di struttura e di evoluzione del manto vegetale, dove, in seguito allo sviluppo secondario di una vegetazione di sponda seguiva, negli anni successivi, la crescita delle specie più propriamente acquatiche. Le cave di Noale e Salzano descritte in letteratura (Anoè, Caniglia, 1987) hanno caratteristiche simili e inoltre Salzano, in particolare, ha subito una serie di trasformazioni per adibirle alla fitodepurazione che la rendono comparabile con l’impianto di Campocroce.
Come nel caso di Salzano anche nell’area studiata si sono sviluppate proprio le specie igrofile che solitamente sono scelte da chi opera nel campo della fitodepurazione.
I risultati di questa ricerca portano a suggerire che nella maggioranza dei casi, la messa a dimora delle specie acquatiche con funzione fitodepurante, possa essere soltanto una anticipazione di quello che la normale evoluzione della vegetazione riesce a compiere nel giro di pochi anni senza l’utilizzo di energie esterne e risorse finanziarie. Tale tipologia di analisi richiede comunque un tempo maggiore di rilevamento e soprattutto una serie di indagini di tipo pedologico e chimico sulle acque che scorrono prima e dopo il filtro vegetale.
Un altro importante aspetto messo in luce con la presente ricerca, è stato attestare il comportamento ecologico di queste fitocenosi, caratterizzate da una grande capacità dinamica di colonizzazione dei suoli inondati.
Appendice Tabella 1 – Altre specie
L’ordinamento segue quello della tabella 1
Se non indicato diversamente, la presenza della specie è da intendersi (+).
Vedi Fig. 2a
Area di rilevamento 1
Erigeron annuus, Abutilon theophrasti, Lolium multiflorum, Alopecurus myosuroides (1), Rumex acetosa, Trifolium pratense, Medicago lupulina (1), Carex panicea, Rumex crispus.
Area di rilevamento 2
Convolvulus arvensis, Arctium lappa, Rumex acetosa, Equisetum telmateja.
Area di rilevamento 3
Galium album, Taraxacum officinale, Paspalum paspaloides.
Area di rilevamento 4
Erigeron annuus, Lolium multiflorum, Arctium lappa, Alopecurus myosuroides, Rorippa sylvestris, Matricaria chamomilla, Tiphoides arundinacea.
Vedi Fig. 2b
Area di rilevamento 5
Erigeron annuus, Abutilon theophrasti, Taraxacum officinale, Rumex crispus, Cynodon dactylon, Festuca arundinacea, Centaurea nigrescens, Hypericum perforatum, Agropyron repens, Paspalum paspaloides, Carex panicea.
Area di rilevamento 12
Cyperus glomeratus, Eupatorium cannabinum, Carex distans.
Area di rilevamento 14
Erigeron annuus, Tamarix gallica, Convolvulus arvensis, Ambrosia artemisiifolia, Equisetum arvense, Juncus inflexus.
Area di rilevamento 13
Trifolium pretense, Ambrosia artemisiifolia, Carex distans.
Area di rilevamento 15
Typhoides arundinacea, Tamarix gallica, Setaria viridis.
Area di rilevamento 16
Erigeron annuus, Lolium multiflorum, Verbena officinalis, Carex panicea.
Area di rilevamento 17
Convolvulus arvensis.
Area di rilevamento 18
Tamarix gallica, Gratiola officinalis, Platanus hybrida, Velezia rigida, Carex distans.
Area di rilevamento 8
Epilobium hirsutum, Prunella vulgaris, Setaria viridis, Cynodon dactylon (+.3), Juncus inflexus, Juncus geradii.
Area di rilevamento 7
Epilobium hirsutum, Juncus inflexus, Juncus gerardii, Galium album, Prunella vulgaris, Rorippa sylvestris, Equisetum telmateja, Crepis biennis, Festuca pratensis, Lotus corniculatus, Verbena officinalis.
Area di rilevamento 6
Tamarix gallica, Carex distans, Abutilon teophrasti, Epilobium hirsutum, Prunella vulgaris, Setaria viridis, Cyperus glomeratus (+3), Blackstonia perfoliata, Carex hirta.
Area di rilevamento 10
Tamarix gallica, Abutilon theophrasti, Epilobium hirsutum, Galium album, Verbena officinalis, Gratiola officinalis, Crepis biennis, Polygonum aviculare, Equisetum palustre, Typhoides arundinacea, Samolus valerandi, Juncus gerardii, Carex panicea, Cyperus esculentum
Area di rilevamento 11
Cuscuta campestris, Samolus valerandi.
Area di rilevamento 9
Arctium lappa, Blackstonia perfoliata, Samolus valerandi, Stachis palustris
Bibliografia
Anoè N., Caniglia G. (1987) – La vegetazione acquatica e palustre di alcune cave di argilla dell’entroterra veneziano. Lavori Soc. Ven. Sc. Nat., 21: 159-175.
Blasi C., Paolella A. (1992) – Progettazione ambientale. Cave, fiumi, strade, parchi, insediamenti. NIS, La Nuova Italia Scientifica.
Borin M., Tocchetto D. (2007) – Five year water and nitrogen balance for a constructed surface flow wetland treating agricultural drainage waters. Sc. Total Envir., 380: 38-47.
Braun Blanquet J. (1928) – Pflanzensoziologie. Springer, Berlin.
Buffa G., Carpenè B., Casarotto N., Da Pozzo M., Filesi L., Lasen C., Marcucci R., Masin R., Prosser F., Tasinazzo S., Villani M., Zanatta K. (2016) – Lista rossa regionale delle piante vascolari della regione del Veneto. Parco Regionale Dolomiti d’Ampezzo, Regione Veneto, Società Botanica Italiana.
Ellenberg H. (1988) – Vegetation Ecology of Central Europe. Cambridge University Press, Avon.
Ellenberg H., Weber H.E., Dull R., Wirth V., Werner W., Pauliben D. (1992) – Indicator values of plants in Central Europe. Scripta geobotanica, 18: 258 pp.
Gerdovic L. (1992) – Depurazione dei sistemi acquatici. Mensile dell’accademia Russa delle scienze, 9 (925): 26-33.
Ispra (2012) – Guida tecnica per la progettazione e realizzazione di impianti di fitodepurazione per il trattamento delle acque reflue urbani. Manuali e Linee Guida 81; 182 pp.
Moroni F., Pergetti M., Ghetti P.F. (1996) – La fitodepurazione con il sistema a flusso subsuperficiale: principi di funzionamento e aspetti progettuali di un impianto per il trattamento di reflui civili. In Atti del Convegno: Fitodepurazione, metodologie ed applicazioni. Finale Emilia: pp 137-146.
Oberdorfer E. (1977, ’78, ’83, ’92) – Suddeutsche Pflanzengesellschaften. Voll. 1, 2, 3, 4,ed. Gustav Fischer Verlag, Stuttgart, New York
Pignatti S. (1982) – Flora d’Italia. Voll. 1,2,3, Ed. Agricole, Bologna.
Pignatti S. (1994) – Ecologia del paesaggio. Utet, Torino: 225 pp.
Pignatti S. (1998) – I boschi d’Italia. Sinecologia e biodiversità. Scienze Ambientali e naturali, UTET, Torino: 67 pp.
Raunkiaer (1934) – The life of plants and statistical plant geography. Oxford
Veneto Agricoltura (2014) – La fitodepurazione per il trattamento di acque di origine agricola e di reflui zootecnici. Edizioni MB srl, 48 pp.
Vismara R. (1998) – Depurazione biologica – Teoria e processi. Hoepli Ed.: 778 pp.
Vymazal, J. (2011) – Plants used in constructed wetlands with horizontal subsurface flow: a review. Hydrobiologia, 674 (1): 133-156.
Vymazal, J. (2013) – Emergent plants used in free water surface constructed wetlands: A review. Ecological Engineering, 61:582-592.
Zangheri P. (1976) – Flora italica. Cedam, Padova, vol. 1 e 2
Siti Web
http://www.prodromo-vegetazione-italia.org/