Riassunto
Viene descritta una fauna di crostacei fossili associati a coralli dell’Eocene inferiore proveniente da accumuli organogeni affioranti nel territorio del comune di Altissimo (Valle del Chiampo – Vicenza) comprendente i rappresentanti di una famiglia di Axiidea, sette di Anomura e diciassette di Brachyura. Sono presentati 75 taxa. I seguenti taxa sono di nuova istituzione: Cristinagalathea striata gen. nov., sp. nov. (Munidopsidae), Vicetitrapezia exaltissimo gen. nov., sp. nov. (Vicetitrapeziidae fam. nov. di Trapezioidea), Clampethildella spinosa gen. nov., sp. nov. (Mathildellidae). È stato inoltre istituito Muellermunida gen. nov. (Munididae) per una migliore collocazione sistematica di Protomunida pentacantha e di Eumunida veronensis. Viene fornita una diagnosi emendata di Paguristes vallionensis (Diogenidae). È presentato un esemplare di Etisus arduinoi con caratteristiche giovanili. L’indice di biodiversità Shannon-Weaver calcolato per questa fauna è conforme a quelli relativi alle faune ypresiane del Veneto di ambiente corallino già note.
Abstract
The coral-associated Crustacean fauna from the early Eocene in the Upper Chiampo Valley (Altissimo – Vicenza – NE Italy)
An early Eocene coral-associated crustacean fauna from bioconstructions cropping out at Altissimo (Chiampo Valley – Vicenza) is described with representatives of one family within the Axiidea, seven within the Anomura and seventeen within the Brachyura. The taxa considered are 75. The following taxa are new: Cristinagalathea striata gen. nov., sp. nov. (Munidopsidae), Vicetitrapezia exaltissimo gen. nov., sp. nov. (Vicetitrapeziidae fam. nov. within the Trapezioidea), Clampethildella spinosa gen. nov., sp. nov. (Mathildellidae). Muellermunida gen. nov. (Munididae) is established for a better accomodation of Protomunida pentacantha and of Eumunida veronensis. An emended diagnosis for Paguristes vallionensis (Diogenidae) is given. One juvenile specimen of Etisus arduinoi is presented. The Shannon-Weaver biodiversity index calculated for this fauna is in accordance with the ones relative to the already known Ypresian faunas from coralligenous buildups in Venetia.
Introduzione
Il presente lavoro fa seguito a ricerche sul campo indirizzate allo studio delle faune a crostacei presenti nelle piccole scogliere ad alghe e coralli dell’Eocene inferiore che emergono in più punti della Valle del Chiampo.
Le prime segnalazioni al riguardo sono relative alla fauna rinvenuta nella cava di contrada Gecchelina di Monte di Malo (Vicenza), cui sono succeduti gli studi su quelle di San Pietro Mussolino (Vicenza) e Vestenanova (Verona) e la presentazione della ricca fauna proveniente da varie località attorno a Bolca (Verona) (Beschin et al., 2000, 2007, 2015, 2016; Tessier et al., 2011).
Sino agli anni 2000 i rinvenimenti di crostacei fossili associati a coralli nel territorio italiano erano assai scarsi, per la difficoltà di riconoscere resti di organismi adattati a vivere all’interno delle colonie di coralli e, pertanto, di dimensioni millimetriche.
Sicuramente tale dato era fortemente influenzato anche dalle modalità con cui si procedeva alle ricerche sul campo, che mettevano in luce con maggior facilità esemplari di dimensioni ragguardevoli, tralasciando i resti fossili di dimensioni spesso ridottissime, che richiedevano tecniche di raccolta diverse (prelievo delle rocce in situ e uso di lenti e stereomicroscopi in laboratorio) (Beschin et al., 2017).
Affinando quindi le metodologie di ricerca ed intensificando le uscite sul territorio, sono stati scoperti nuovi siti che si sono rivelati assai ricchi di reperti riferibili all’Eocene inferiore.
I risultati di queste ricerche hanno messo in luce l’incredibile abbondanza e varietà delle faune fossili, permettendo anche di effettuare un’analisi comparativa che ha dimostrato la presenza di un alto indice di biodiversità, analogo a quello delle faune attuali che popolano le acque basse delle scogliere coralline del Pacifico Centrale (Beschin et al., 2017).
E questi studi hanno dato il via ad ulteriori approfondimenti sempre rivolti alle faune a crostacei dell’Eocene inferiore del Veneto (De Angeli & Ceccon, 2012, 2013a,b, 2014, 2015, 2016a,b, 2017, 2020; De Angeli & Garassino, 2014; De Angeli & Caporiondo, 2016, 2020).
Nel presente lavoro sono presi in considerazione i resti di decapodi provenienti dagli affioramenti individuati nella parte medio-alta della Valle del Chiampo, in sinistra idrografica del torrente omonimo nel comune di Altissimo (Vicenza). Sono stati analizzati 344 esemplari, che hanno permesso di riconoscere 25 famiglie appartenenti a tre sottordini: Axiidea de Saint Laurent, 1979, Anomura Milne-Edwards, 1832 e Brachyura Linnaeus, 1758. All’interno dei 75 taxa riconosciuti sono stati definiti una nuova famiglia di Trapezioidea Miers, 1886, quattro nuovi generi e tre nuove specie.
Aspetti geografici e geologici del territorio di Altissimo
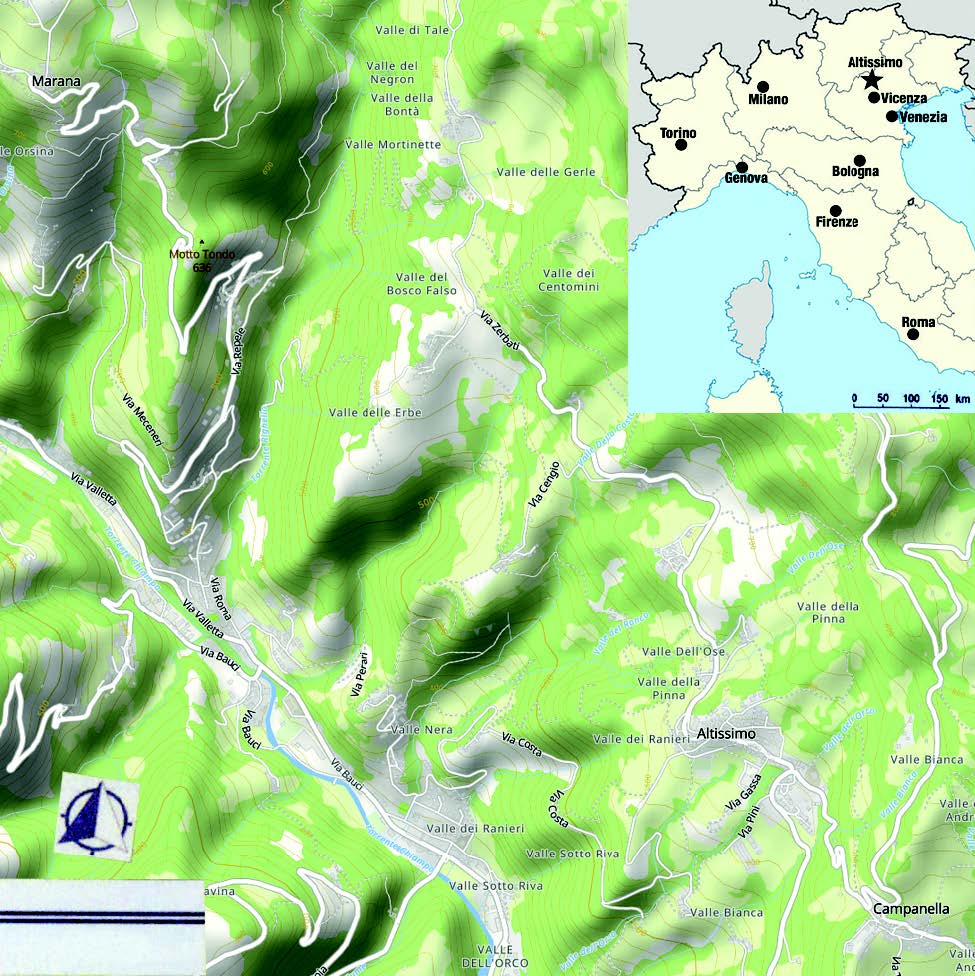
Altissimo è un comune del Veneto in provincia di Vicenza adagiato ad oltre 600 m di altitudine sui contrafforti sudorientali dei Monti Lessini. Il suo territorio, che fa parte del Parco Naturale Regionale della Lessinia e si estende per circa 15 km², ha configurazione poligonale: più stretto nella parte settentrionale, si espande notevolmente e con regolarità procedendo verso Sud. Longitudinalmente esso è diviso in due parti dal torrente Chiampo: la parte orientale, sulla sinistra idrografica, accoglie gli affioramenti indagati e anche il centro abitato sede del Municipio; la parte poco estesa che rimane a Ovest, sulla destra della valle, raggiunge Monte Postale di Bolca, ai confini della provincia di Verona (fig. 1). È un territorio suggestivo e turistico che da quote piuttosto basse sul fondovalle (mediamente 300 m s.l.m.) si eleva notevolmente nell’area estrema settentrionale del comune, addossata a Cima Marana (circa 1350 m s.l.m.).
Nell’area d’indagine affiorano prevalentemente formazioni sedimentarie marine di età compresa tra il Cretaceo inferiore e l’Eocene medio e rocce vulcaniche. Queste ultime possono essere di tipo effusivo o piroclastico (basalti, tufi, ialoclastiti, brecce e vulcanoclastiti) e si legano all’intenso magmatismo che caratterizzò il Paleogene delle Venezie in più episodi (Piccoli, 1966). Come noto, l’attività vulcanica, qui come in tutti i Lessini orientali e i Berici occidentali, fu condizionata dalla presenza di una fossa tettonica di tipo distensivo molto subsidente, detta graben dell’Alpone-Chiampo, delimitata ad Ovest dalla faglia di Castelvero con la sua corte di faglie sub-parallele orientate NNO-SSE, che permise la deposizione di grandi spessori di materiali vulcanici (Mietto, 1997; Beschin et al., 2016). Le rocce piroclastiche, diffuse ma non particolarmente resistenti agli agenti atmosferici, danno luogo ad un paesaggio dall’aspetto dolce. Fanno eccezione alcuni rilievi molto ripidi di forma conica, che rappresentano i riempimenti a composizione basaltica di antichi condotti vulcanici messi par
zialmente a giorno dagli agenti esogeni; si tratta dei neck della Bressavalda, di contrada Zerbati e di Campanella.
Le rocce sedimentarie marine più antiche del territorio di Altissimo appartengono al Mesozoico e sono costituite da un piccolo affioramento di Calcari Grigi del Giurassico inferiore che si osserva ai limiti settentrionali del comune. Notevolmente più sviluppati sono, invece, le stratificazioni del Cretaceo inferiore (particolarmente la Maiolica) e quelli del Cretaceo superiore (Scaglia Rossa) che si osservano nella zona valliva sulle due sponde del torrente. La Maiolica ha spessore variabile, ma comunque attorno ai 100 m, ed è formata da calcari di prevalente origine biochimica indicativi di condizioni marine abbastanza profonde. La Scaglia Rossa invece è una formazione meno potente e di ambiente solo relativamente profondo, più ossigenato.
È noto che praticamente in tutto il Vicentino, al di sopra della Scaglia Rossa, è registrata una lacuna deposizionale comprendente buona parte del Paleocene (Mietto, 1997; Beschin et al., 2016); il dato è confermato anche per l’area in esame. Durante le fasi di quiete del successivo vulcanesimo eocenico, nel graben dell’Alpone-Chiampo si depositarono materiali carbonatici molto ricchi di fossili tipici di un ambiente di piattaforma continentale con acque poco profonde, calde e agitate (De Zanche, 1965; Beschin et al., 1991). Per l’Eocene inferiore si tratta di calcari di scogliera ad alghe e coralli, eteropici alle vulcaniti, da cui provengono i resti di crostacei studiati e di calcari nummulitici stratificati che per le caratteristiche litologiche e paleontologiche e a volte per la continuità laterale sono correlabili con gli analoghi affioramenti dell’area di Bolca, Vestenanova (Cava Braggi) e San Pietro Mussolino (Cava Bertocchi) (Tessier et al., 2011;
Beschin et al., 2015, 2016) e quindi vanno ascritti allo Ypresiano. Ridottissime sono le assise di calcari dell’Eocene medio: affiorano presso la contrada Santolini e presso la chiesa di Altissimo; di esse in passato si è localmente tentato anche lo sfruttamento nell’industria del marmo, ma con scarsi risultati per l’impurità della roccia.
Dal punto di vista tettonico, questa parte di Lessini centro-orientali si presenta come una estesa monoclinale immergente a Sud, variamente interessata da faglie lungo le quali i corsi d’acqua e in particolare il torrente Chiampo hanno operato la loro azione erosiva e modellato il territorio nell’attuale aspetto. Nella parte meridionale dell’area considerata e prossima a San Pietro Mussolino, è presente inoltre una piega che interessa banchi di calcare del Cretaceo. Non va infine sottaciuta la presenza nella parte settentrionale del territorio dell’importante “faglia pedemontana”, che si pone sulla prosecuzione occidentale della piega a ginocchio che delimita verso Sud le Prealpi venete (Visonà, 1974; Cocco, 1975; Cocco & Boscardin, 2004).
Le formazioni dello Ypresiano del versante vicentino del Monte Postale (Altissimo), che si trova a brevissima distanza dalla famosa “Pesciara”, sono state trattate in un lavoro specifico dedicato all’intera area di Bolca, al quale si rimanda (Beschin et al., 2016).
Materiali e metodi
Ricerca geologica sul campo
Sono state effettuate da parte di uno degli autori (CB) ricerche sistematiche nel territorio del comune di Altissimo per individuare biocostruzioni riferibili allo Ypresiano analoghe a quelle coeve recentemente rinvenute in varie località del Vicentino e del Veronese che hanno permesso di scoprire per quest’area l’esistenza di una vasta laguna eocenica caratterizzata da patch reef (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); i nuovi ritrovamenti consentono di valutarne più precisamente l’estensione.
I crostacei
La fauna di crostacei qui analizzata è costituita da 344 reperti raccolti negli ammassi organogeni affioranti in varie località del comune di Altissimo (Vicenza). Gli esemplari sono in deposito presso il Museo Civico “G. Zannato” di Montecchio Maggiore (Vicenza) e sono indicati con il loro numero di catalogo (MCZ).
I resti sono costituiti essenzialmente da carapaci e parti di chele di dimensioni molto piccole e si presentano in buono stato di conservazione. La raccolta è stata effettuata prelevando dagli affioramenti dei campioni di roccia. In laboratorio il materiale è stato ridotto in frammenti più piccoli permettendo il recupero e la successiva preparazione di tutti i resti di crostacei.
Il lavoro di campionamento è stato interrotto quando i risultati già conseguiti in termini di composizione della fauna e di abbondanza relativa dei taxa risultavano ormai definiti.
Il materiale è stato preparato con l’aiuto di una lente di ingrandimento con lampada scialitica e di un microscopio stereoscopico. Tutti gli esemplari sono stati misurati con un calibro digitale. Le dimensioni sono espresse in millimetri. Nel testo si farà riferimento ai seguenti parametri biometrici (figg. 2a,b):
– L: larghezza massima del carapace;
– l: lunghezza massima del carapace (compreso il rostro);
– Lo: ampiezza del margine fronto-orbitale;
– lpr: lunghezza massima del propodo;
– lp: lunghezza del palmo;
– h: altezza massima del palmo.
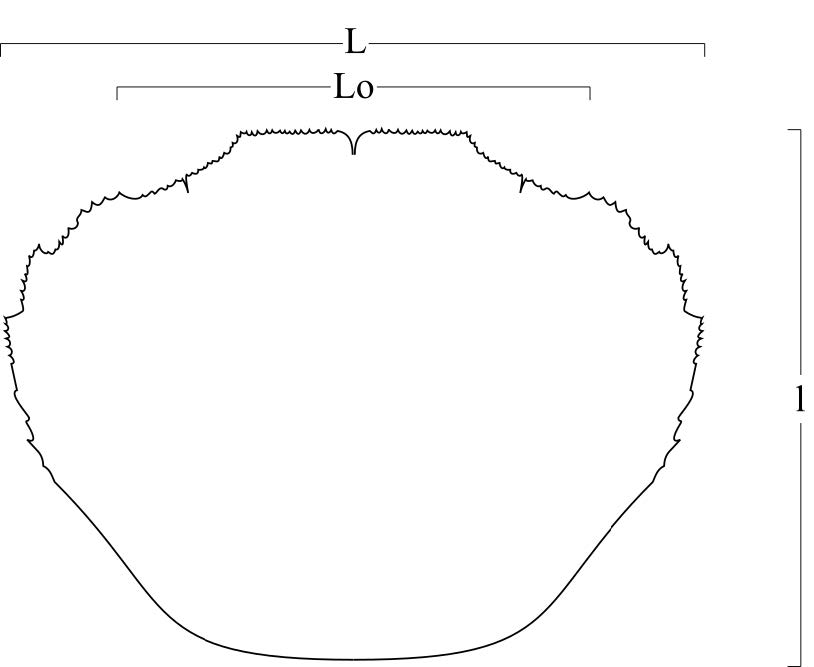
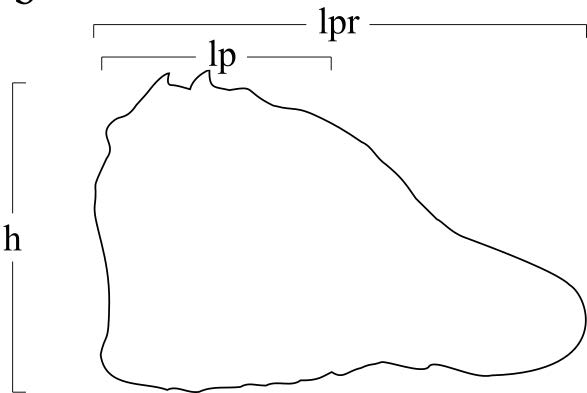
Fig. 2. Schema delle misurazioni. a: carapace, b. propodo di chelipede.
Per le specie di nuova istituzione è stata fornita una diagnosi in italiano e una in inglese, la descrizione e alcune osservazioni con gli opportuni confronti. Per le specie già note è stato fornito il riferimento al lavoro istitutivo, per le citazioni intermedie il riferimento al catalogo De Angeli & Garassino (2006) e quindi le citazioni successive ad esso; per le specie non presenti in questo catalogo è stato riportato il riferimento ad un eventuale altro lavoro comprendente citazioni intermedie e le citazioni successive. In ogni caso sono state riportate solo le citazioni che si riferiscono a lavori relativi a nuovi ritrovamenti dei taxa o a modifiche della loro posizione sistematica. Ove necessario è stata inserita una descrizione a integrazione di quella originale e/o delle osservazioni.
Per l’inquadrameto sistematico si è fatto riferimento ad Ahyonget al. (2010), Schweitzer et al. (2010, 2012, 2018, 2020). Si è riconosciuta la validità della Superfamiglia Trapezioidea Miers, 1886 come proposto da Nget al. (2008). Nell’elenco sistematico si è preferito non utilizzare il rango sottofamiglia.
Risultati
Ammassi organogeni
Sono stati individuati ammassi organogeni in precedenza non noti nelle località di seguito elencate dove affiorano piccole scogliere chiaramente eteropiche alle vulcaniti e localmente associate ai Calcari Nummulitici; da Nord a Sud: contrada Cocco (CC), contrada Zerbati (CZ), via Cero (VC), Valle dei Pini (VdP) e via Pezzi (VP) (fig. 1).
Come già accennato, le biocostruzioni si presentano in forme di ammassi sparsi tra le rocce vulcaniche eoceniche costituite da ialoclastiti basaltiche, basalti compatti, camini con brecce, ecc. La loro entità è in genere modesta. Va rilevato che nell’area settentrionale del territorio considerato (contrade Cocco e Zerbati), tali emergenze sono sporadiche tra le superfici a prato o a coltivo (fig. 3). Più in evidenza, invece, l’accumulo di via Cero che si osserva a lato della strada che congiunge la frazione di Molino ad Altissimo, prima di incontrare il cimitero; si tratta di calcari molto cristallizzati di colore bianco-giallastro in facies chiaramente di scogliera (fig. 4). Gli affioramenti di Valle dei Pini sono costituiti da calcari cristallini di colore biancastro poco compatti con abbondantissime alghe calcaree. Un cenno particolare merita, infine, il sito di Via Pezzi che è stato messo di recente allo scoperto da lavori di scavo di allargamento della via nell’incontro con la strada provinciale 44 che collega San Pietro Mussolino a Campanella (fig. 5). Qui si osserva un consistente lembo di calcari biancastri e giallo-rossicci più o meno cristallizzati, parzialmente in facies di scogliera, che fa passaggio verso Nord a brecce e rocce basaltiche compatte. A contrada Cocco sono stati raccolti 22 esemplari, a contrada Zerbati 48, in via Cero 83, in Valle dei Pini 102, in via Pezzi 89.
Oltre alla fauna di crostacei di seguito studiata, i bioherma hanno restituito resti di bivalvi (fig. 6), modelli interni di gasteropodi in alcuni casi fortemente cristallizzati (fig. 7) e anche minuscoli echinidi (fig. 8), oltre che abbondanti alghe e coralli (fig. 9).
Analisi sistematica della fauna di crostacei
Ordine Decapoda Latreille, 1802
Infraordine Axiidea de Saint Laurent, 1979
Famiglia Callichiridae Manning & Felder, 1991
Genere Neocallichirus K. Sakai, 1988
Specie tipo: Neocallichirus horneri K. Sakai, 1988
Neocallichirus sp.
(fig. 10)
Materiale e dimensioni. Due propodi di chelipede destro (minore) in visione interna: (VdP) MCZ 5878 lpr: 7,3 lp: 4,1 h: 2,5; MCZ 5879 h: 3,7.
Descrizione. Propodo minore nel complesso piuttosto snello. Palmo rettangolare allungato con margini superiore e inferiore poco convergenti distalmente e con angoli prossimali convessi; presso il margine prossimale, che si presenta subverticale, si osserva un’ampia concavità legata all’articolazione con il carpo. Margine superiore appena convesso; margine inferiore nel primo tratto convesso, concavo alla base del dito fisso. Margine distale regolarmente concavo; a livello dell’articolazione con il dattilo si osservano quattro minuscoli denti appuntiti di dimensioni decrescenti distalmente; piccoli denti si osservano anche nella parte prossimale del margine occlusale del dito fisso. Dito fisso sottile e appuntito, quasi diritto, poco più corto del palmo. Lungo il margine inferiore di tutto il propodo si osservano nove piccole spine appuntite rivolte in avanti, otto ravvicinate a livello del palmo e della base del dito fisso, la nona isolata circa a metà del dito fisso. Superficie interna del palmo molto bombata in senso longitudinale e trasversale.
Osservazioni. La classificazione dei resti di chele fossili di crostacei ed in particolare di Axiidea è sempre molto problematica, soprattutto quando si presentano incompleti, come nel caso del reperto di Altissimo, di cui è conservato unicamente il propodo.
Per la forma generale esso ricorda quello delle chele minori di alcune specie di Neocallichirus, genere, probabilmente originato nella Tetide, diffuso oggi nell’Indo-Pacifico e nell’Atlantico occidentale; le numerose specie attribuite al taxon caratterizzano con la loro presenza la zona litorale e la piattaforma interna dove si scavano delle tane ramificate con due aperture.
La diagnosi istitutiva del genere non si sofferma però sulla forma dei chelipedi, ma sulle caratteristiche del carapace e sulla struttura del 3° massillipede; Sakai (1988) sottolinea la vicinanza di Neocallichirus a Callichirus Stimpson, 1866, Glypturus Stimpson, 1866 e Calliax de Saint Laurent, 1973, e indica come carattere diagnostico del genere soprattutto la forma equidimensionale del telson, lungo quanto l’uropodo il cui endopodo si allarga posteriormente.
Anche Schweitzer & Feldmann (2002) e Schweitzer et al. (2004, 2005) trattano di Neocallichirus e sottolineano la difficoltà di attribuire al genere resti fossili, in particolare per la confusione esistente in letteratura dovuta alla scarsa definizione dei caratteri diagnostici dei chelipedi maggiori, i più facilmente conservabili. Gli autori hanno quindi cercato di individuarne alcuni; tra questi: carpo usualmente non rettangolare con margine inferiore arrotondato, articolazione carpo-propodiale perpendicolare all’asse del propodo, palmo rettangolare con un rigonfiamento dentellato più o meno pronunciato sul margine distale, dito fisso relativamente sottile e tendenzialmente privo di denti che si prolunga quasi diritto dal palmo, dattilo robusto ed armato con un dente nella parte prossimale; ritengono comunque che in presenza del solo propodo l’attribuzione non può che essere incerta, attesa la sua morfologia variabile, mentre una classificazione più sicura può essere fatta solo in presenza di altri segmenti del chelipede maggiore.

Fig. 4. Ammasso organogeno affiorante a via Cero.
Fig. 5. Ammasso organogeno di via Pezzi.
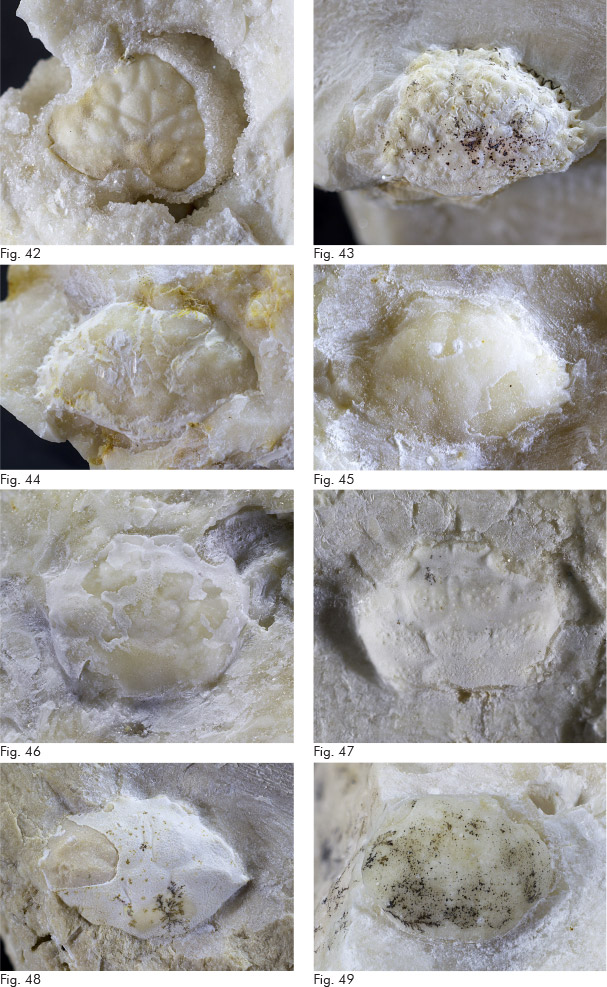
Fig. 6. Piccolo bivalve MCZ 6223 (L: 7,9 l: 6,8); via Cero.
Fig. 7. Modello interno di gasteropode MCZ 6224 (diametro base: 4,0); Valle dei Pini.
Fig. 8. Echinide MCZ 6222 (h: 3,3); contrada Zerbati.
Fig. 9. Ammasso corallino MCZ 5992.
Anche Hyžný & Karasawa (2012) hanno affrontato il problema della classificazione dei resti di chelipedi, trattando in particolare dei generi Neocallichirus, Sergio Manning & Lemaitre, 1994, Podocallichirus K. Sakai, 1999 e Grynaminna Poore, 2000 e ribadendo la necessità per i paleontologi di poter fruire di caratteri diagnostici legati a queste parti dell’organismo e ipotizzando più metodologie di approccio alla materia. Trattando nello specifico di Neocallichirus, questi autori elencano le specie fossili all’epoca inserite nel genere e, convinti della necessità di un lavoro di revisione, esprimono dubbi sulla corretta attribuzione di buona parte delle stesse, senza peraltro fornire argomentazioni a supporto della critica; concludono la loro analisi rilevando che per una corretta classificazione delle chele dei “callianassidi” è necessario poter disporre di tutti i segmenti del chelipede (ischio, mero, carpo, propodo e dattilo) e che ulteriori studi sono necessari sui rappresentanti attuali del gruppo per meglio definirne i caratteri diagnostici.
Ferme queste problematiche, il taxon è ben documentato nel Paleogene del Veneto; per l’Eocene inferiore è segnalato Neocallichirus allegranzii Beschin, De Angeli, Checchi & Zarantonello, 2005 di Monte Serea (Verona) rinvenuto anche in livelli dell’Eocene medio a Cava Grola di Cornedo Vicentino (Vicenza) (Beschin et al., 2005, 2009b, 2012); dell’Eocene medio è anche N. fortisi Beschin, Busulini, De Angeli & Tessier, 2002 di Cava Main di Arzignano e Cava Grola di Cornedo Vicentino (Vicenza) (Beschin et al., 2002, 2012); da livelli dell’Eocene superiore provengono Neocallichirus sp. di Beschin et al. (2006) di Priabona (Vicenza), come pure N. bacatus Busulini, Beschin & Tessier, 2012 e Neocallichirus sp. di Busulini et al. (2012) di Possagno. De Angeli & Beschin (2008) hanno segalato Neocallichirus cfr. N. allegranzii dell’Oligocene inferiore dei Monti Berici (Vicenza).
Hyžný & Zorn (2020) considerano Neocallichirus borensis Beschin, De Angeli, Checchi & Mietto, 2006 dell’Eocene superiore di Priabona (Vicenza) sinonimo di Calianassa tuberculata (sic!) Lőrenthey & Beurlen, 1929 (Eocene medio, Ungheria) ora riferita al genere attuale Karumballichirus Poore, Dworschak, Robles, Mantelatto & Felder, 2019 (Lőrenthey & Beurlen, 1929; Beschin et al., 2006).
L’attribuzione al genere di tutte le specie note per l’Eocene italiano è basata su chelipedi maggiori; l’unico chelipede minore rinvenuto è riferito a N. fortisi (cava Main di Arzignano) (Beschin et al., 2002) che però ha caratteristiche diverse da quelle dell’esemplare qui studiato. Affinità si riscontrano con Neocallichirus sp. di Beschin et al. (2006) che ha analogo profilo sinuoso del margine inferiore del propodo, che però è caratterizzato da una fine denticolazione anziché da alcune evidenti spine. Somiglianze si osservano anche con la chela minore dell’attuale N. raymanningi Blanco-Rambla & Lemaitre, 1999 che tuttavia non possiede le pronunciate spine sul margine inferiore evidenti nell’esemplare fossile, bensì ciuffi di sete (Blanco-Rambla & Lemaitre, 1999). Viste la difficoltà riferite si è preferito lasciare indeterminata l’attribuzione specifica del propodo studiato.
Infraordine Anomura MacLeay, 1838
Superfamiglia Galatheoidea Samouelle, 1819
Famiglia Galatheidae Samouelle, 1819
Genere Acanthogalathea Müller & Collins, 1991
Specie tipo: Galathea (Acanthogalathea) parva Müller & Collins, 1991
Acanthogalathea broglioi Beschin, Busulini & Tessier, 2016
(fig. 11)
2016 Acanthogalathea broglioi Beschin, Busulini & Tessier in Beschin et al., p. 25, fig. 11, t. 1, ff. 2a,b.
Materiale e dimensioni. Due carapaci: (CZ) MCZ 5880 L: 3,7; (VC) MCZ 5881 L: 3,8 l: 6,0.
Distribuzione. Eocene inferiore: Veneto (Rama di Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Acanthogalathea devecchii Beschin, Busulini & Tessier, 2016
(fig. 12)
2016 Acanthogalathea devecchii Beschin, Busulini & Tessier in Beschin et al., p. 26, fig. 12, t. 1, ff. 3a,b, 4.
Materiale e dimensioni. Una porzione di carapace che conserva solo la regione cardiaca e parte di quella branchiale sinistra: (VdP) MCZ 5882 L > 9,1.
Osservazioni. Nonostante la sua lacunosità il reperto viene attribuito alla specie in quanto sono visibili alcuni tra i caratteri diagnostici più significativi e precisamente: margine laterale moderatamente convesso con alcune spine, regione cardiaca con una serie trasversale di tre tubercoli seguita da strie di dimensione decrescente, regioni branchiali scarsamente striate con tre spine mesobranchiali e cinque metabranchiali.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Genere Bolcagalathea Beschin, Busulini & Tessier, 2016
Specie tipo: Bolcagalathea corallina Beschin, Busulini & Tessier, 2016
Bolcagalathea corallina Beschin, Busulini & Tessier, 2016
(fig. 13)
2016 Bolcagalathea corallina Beschin, Busulini & Tessier in Beschin et al., p. 29, fig. 14, t. 1, f. 6, t. 2, ff. 1a, b.
2017 Bolcagalathea corallina – De Angeli & Ceccon, p. 9, t. 2, ff. 1, 2.
Materiale e dimensioni. Tre carapaci: (VC) MCZ 5884 L: 2,8; MCZ 5883 L: 4,2; MCZ 5885.
Distribuzione. Eocene inferiore: Veneto (Rama di Bolca – Verona; Monte Magrè di Schio, Altissimo – Vicenza) (Beschin et al., 2016; De Angeli & Ceccon, 2017).
Bolcagalathea multispinosa Beschin, Busulini & Tessier, 2016
(fig. 14)
2016 Bolcagalathea multispinosa Beschin, Busulini & Tessier in Beschin et al., p. 31, fig. 15, t. 2, ff. 2a, b, 3.
Materiale e dimensioni. Due carapaci incompleti: (CC) MCZ 5887 L > 4,3; (VP) MCZ 5886.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Genere Lessinigalathea De Angeli & Garassino, 2002
Specie tipo: Lessinigalathea regalis De Angeli & Garassino, 2002
Lessinigalathea regalis De Angeli & Garassino, 2002
(fig. 15)
2000 Galathea sp. – Beschin et al., t. 1, f. 4.
2002 Lessinigalathea regale De Angeli & Garassino, p. 13, fig.10, t. 4, f. 1.
2016 Lessinigalathea regalis – Beschin et al., p. 34, t. 2, f. 6 (cum syn.).
2017 Lessinigalathea regalis – De Angeli & Ceccon, p. 11, fig. 4; t. 1, ff. 5-6.
Materiale e dimensioni. Tre carapaci incompleti: (VdP) MCZ 5888 L: 7,8; MCZ 5889; MCZ 5890.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Vestenanova, Bolca – Verona) (De Angeli & Garassino, 2002; Beschin et al., 2007, 2015, 2016; Tessier et al., 2011; De Angeli & Ceccon, 2017).
Lessinigalathea cfr. regalis De Angeli & Garassino, 2002
Materiale e dimensioni. Due carapaci molti frammentari: (VC) MCZ 5892 L: 5,4; (VdP) MCZ 5891.
Osservazioni. L’incompletezza dei resti non consente una loro sicura attribuzione, ma per le loro carattestiche risultano molto vicini a L. regalis.
Genere Palaeomunida Lőrenthey, 1902
Specie tipo: Palaeomunida defecta Lőrenthey, 1902
Palaeomunida sp.
(fig. 16)
Materiale e dimensioni. Due propodi di chelipede in visione interna: (CZ) MCZ 5912 h: 2,2 (destro); MCZ 5913 h: 2,3 (sinistro).
Descrizione. Propodi destro e sinistro simili. Palmo allungato di forma rettangolare con margini superiore ed inferiore subparalleli e margine distale obliquo con piccola convessità sotto l’inserzione del dattilo. Quattro spine dirette in avanti lungo il margine superiore. Dito fisso abbastanza lungo e rastremato. Superficie interna ornata da piccoli tubercoli disposti in allineamenti trasversali che interessano anche il margine inferiore.

Fig. 11. Acanthogalathea broglioi Beschin, Busulini & Tessier, 2016, es. MCZ 5881 (L: 3,8 l: 6,0), carapace: visione dorsale.
Fig. 12. Acanthogalathea devecchii Beschin, Busulini & Tessier, 2016, es. MCZ 5882 (misura parziale L: 9,1), carapace: visione dorsale.
Fig. 13. Bolcagalathea corallina Beschin, Busulini & Tessier, 2016, es. MCZ 5884 (L: 2,8), carapace: visione dorsale.
Fig. 14. Bolcagalathea multispinosa Beschin, Busulini & Tessier, 2016, es. MCZ 5887 (misura parziale L: 4,3), carapace: visione dorsale.
Fig. 15. Lessinigalathea regalis De Angeli & Garassino, 2002, es. MCZ 5888 (L: 7,8), carapace: visione dorsale.
Fig. 16. Palaeomunida sp., es. MCZ 5913 (h: 2,3), propodo di chelipede sinistro: visione interna.
Fig. 17. Eosadayoshia bolcensis Beschin, Busulini & Tessier, 2016, es. MCZ 5893 (L: 3,2 l: 5,0), carapace: visione dorsale.
Osservazioni. Gli esemplari sono molto simili all’esemplare Palaeomunida sp. descritto per Parona (Verona) da Beschin et al. (2019). Trattandosi di esemplari incompleti non è possibile procedere ad una assegnazione specifica.
Famiglia Munididae Ahyong, Baba, Macpherson & Poore, 2010
Genere Eosadayoshia Beschin, Busulini & Tessier, 2016
Specie tipo: Eosadayoshia bolcensis Beschin, Busulini & Tessier, 2016
Eosadayoshia bolcensis Beschin, Busulini & Tessier, 2016
(fig. 17 )
2016 Eosadayoshia bolcensis Beschin, Busulini & Tessier in Beschin et al., p. 35, fig. 17, t. 3, ff. 1, 2 a-c.
Materiale e dimensioni. Due carapaci: (VP) MCZ 5894 L: 2,6; MCZ 5893 L: 3,2 l: 5,0.
Distribuzione. Eocene inferiore: Veneto (Rama di Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Genere Muellermunida gen. nov.
Specie tipo: Protomunida pentacantha Müller & Collins, 1991
Origine del nome: il taxon, inserito per le caratteristiche del rostro tra i Munididae, è dedicato a Pál Müller, che ha contribuito in modo rilevante alla conoscenza delle faune europee di crostacei fossili e che ha istituito la specie tipo del nuovo genere.
Diagnosi. Carapace subquadrato, poco più lungo che largo (escluso il rostro), convesso in senso trasversale. Rostro con base larga, relativamente corto, triangolare, con lunga spina mediana e due spine laterali; la mediale molto sviluppata, separata da un solco da quella laterale; spina extraorbitale sviluppata. Solchi cervicale e branchiali incisi. Margini laterali con sette/otto spine (compresa la spina extraorbitale). Margine posteriore concavo e fortemente bordato. Lobo epigastrico grande e rigonfio con alcune piccole spine. Processo anteriore mesogastrico evidente. Regioni dorsali con poche piccole spine e creste trasversali subparallele alcune interrotte.
Diagnosis. Carapace subsquared, weakly longer than wide (excluding rostrum), transversally convex. Rostrum with broad basis, relatively short, triangular with long median spine and two lateral spines: medial one developed, separated from lateral one by a furrow; outer orbital spine developed. Cervical and branchial grooves sunk. Lateral margins with seven/eight spines (including outer orbital one). Posterior margin concave and strongly rimmed. Epigastric lobe large, swollen with a few tiny spines. Anterior mesogastric process apparent. Dorsal regions with few small spines and transverse subparallel cristae, some of them discontinuous.
Osservazioni. L’attribuzione generica di Protomunida pentacantha Müller & Collins, 1991 dell’Eocene ungherese e italiano (Müller & Collins, 1991; Beschin et al., 2019) è stata riveduta più volte. Schweitzer & Feldmann (2000) avevano ritenuto infatti che la presenza di cinque spine rostrali rendesse più corretto attribuire tale specie a Eumunida Schmitt, 1883, a loro dire l’unico genere di galateoidea con tale caratteristica (scelta che è stata seguita anche da Beschin et al., 2019).
Schnabel & Ahyong(2010) invece, sulla base della conformazione delle spine frontali, la presenza di una fila di spine epigastriche e di alcune spine post-cervicali sul dorso del carapace, hanno attribuito il taxon a Sadayoshia Baba, 1969, genere che conta alcune specie attuali tutte dell’Indo-Pacifico che vivono a una profondità compresa tra 100 e 500 m.
Va tuttavia rilevato che Müller & Collins(1991), pur evidenziando la somiglianza di Protomunida pentacantha con Sadayoshia acroporae Baba, 1972 proprio sulla base dei caratteri citati, sottolineavano come differenza fondamentale la presenza di un rilievo post-frontale a forma di A allargata. Oltre alla citata caratteristica, in P. pentacantha come pure in Eumunida veronensis Beschin, Busulini & Tessier, 2019, rispetto a Sadayoshia il carapace risulta proporzionalmente più allungato ed è differente la conformazione delle spine laterali rostrali: nelle specie citate la prima è particolarmente grande ed è separata dalla seconda da un solco che si continua posteriormente, mentre in Sadayoshia è separata da un seno evidente; i lobi epigastrici appaiono come detto più sviluppati e rigonfi, separati dalle vestigia del processo anteriore mesogastrico; le spine laterali epatiche sono più arretrate, risultando diverso l’andamento del margine sovraorbitale che nelle varie specie di Sadayoshia mostra uno sviluppo orizzontale accentuato. Alla luce di queste caratteristiche Protomunida pentacantha come pure Eumunida veronensis vanno attribuite ad un nuovo, diverso, genere.
Il genere Eosadayoshia (Beschin et al., 2016), pur avendo tra le sue caratteristiche diagnostiche la presenza di un rostro carenato ad apice trifido e spina sopraorbitale analogo a quello delle specie attuali di Sadayoshia Baba, 1969 e anche di Muellermunida gen. nov., si distingue però da entrambi i generi per il rostro più piatto e slanciato con diverso raccordo con la fronte e per la linea trasversale postcervicale presente solo nella parte mediana del dorso.
Muellermunida veronensis (Beschin, Busulini & Tessier, 2019) (comb. nov.)
(fig. 18)
2019 Eumunida veronensis Beschin, Busulini & Tessier in Beschin et al., p. 75, figg. 10, 11.
Materiale e dimensioni. Un carapace: (VC) MCZ 5895 L: 3,8.
Osservazioni. L’esemplare è incompleto ma consente di osservare le basi delle spine epigastriche che sono mal conservate.
Distribuzione. Eocene inferiore: Veneto (Altissimo – Vicenza); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Famiglia Munidopsidae Ortmann, 1898
Genere Cristinagalathea gen. nov.
Specie tipo: Cristinagalathea striata sp. nov.
Origine del nome: Cristinagalathea (f.); il nuovo genere è dedicato alla studiosa Cristina M. Robins (USA) che negli ultimi anni ha contribuito con numerosi lavori alla conoscenza dei Galatheoidea fossili.
Diagnosi. La stessa della specie tipo.
Diagnosis. As the type species.
Cristinagalathea striata sp. nov.
(figg. 19, 20)
Serie tipica. Olotipo: esemplare MCZ 5906 (fig. 20). Paratipi: esemplare MCZ 5905, MCZ 5907, MCZ 5908, MCZ 5909, MCZ 5910, MCZ 5911.
Località tipo: Altissimo, Vicenza
Livello tipo: Ypresiano (Eocene inferiore).
Origine del nome: striata (lat.) = striata; con riferimento alle coste trasversali che ornano la parte posteriore del carapace.
Materiale e dimensioni. Sette carapaci: (VC) MCZ 5906 L: 2,6; MCZ 5905; (VdP) MCZ 5910 L: 2,0 l > 2,6; MCZ 5908 L: 2,6 l: 3,9; MCZ 5909 L: 2,6; MCZ 5907 L: 3,6; MCZ 5911. MCZ 5905 associato a Titanocarcinus raulinianus MCZ 5990; MCZ 5907 associato a Paratetralia convexa MCZ 6114; MCZ 5910 e MCZ 5911 associati a Dromiopsis sp. MCZ 5970 e Prealpicarcinus cfr. laisensis MCZ 6015.
Diagnosi. Carapace più lungo che largo, convesso in senso trasversale. Rostro molto grande, spatolato con apice triangolare e spine marginali. Margini laterali subparalleli con sei spine (esclusa quella extraorbitale). Margine posteriore leggermente concavo e fortemente bordato. Solco circumgastrico inciso. Regione gastrica con nodo epigastrico e cinque brevi creste convesse. Parte posteriore del carapace con cinque creste trasversali ondulate, la terza e la quinta incomplete.
Diagnosis. Carapace longer than wide, transversally convex. Rostrum very large, spatulate, with triangular apex and marginal spines. Lateral margins sub-parallel with six spines (excluding outer orbital one). Posterior margin weakly concave with strong rim. Circumgastric groove sunk. Gastric region with epigastric node and five short convex cristae. Posterior part of carapace with five wavy transverse cristae, third and fifth one incomplete.
Descrizione. Carapace più lungo che largo, convesso soprattutto in sezione trasversale, con massima larghezza circa a metà. Rostro molto sviluppato, largo, spatolato, con superficie interessata da striature longitudinali; i suoi margini sono per un buon tratto quasi paralleli, convergono poi bruscamente disegnando un apice triangolare con vertice sviluppato e appuntito. I margini rostrali sono caratterizzati da piccole spine soprattutto nei tratti convergenti a lato della spina apicale; quella di dimensione maggiore segna la base della porzione triangolare; nei tratti subparalleli si osservano altre due spine, quella laterale è la intraorbitale. Il margine sopraorbitale è nettamente concavo, liscio, delimitato da una spina extraorbitale molto sviluppata. I margini laterali del carapace sono piuttosto lunghi e quasi paralleli, convessi nel tratto posteriore; portano sei spine triangolari dirette lateralmente (esclusa l’extraorbitale): una sul margine epatico, due sul margine epibranchiale e tre sul margine branchiale posteriore; il margine posteriore è ampio, leggermente concavo e delimitato per tutta la lunghezza da un bordino rilevato. La regione gastrica è chiaramente delimitata dal solco circumgastrico e porta: due rilievi ovali epigastrici, tre brevi creste convesse affiancate, di cui quella mediana in corrispondenza della base del processo anteriore mesogastrico e le altre due sui lobi protogastrici, e infine due creste convesse appaiate a livello del corpo del lobo mesogastrico. Le regioni epatiche sono piccole, lisce e nettamente delimitate dai rami del solco cervicale; i lobi epibranchiali sono subtriangolari, lisci, ben definiti dai solchi branchiali che hanno andamento
trasversali e non è suddivisa in regioni da solchi; essa è delimitata anteriormente da una prima cresta che segue al centro la parte mediana concava del solco cervicale e ai lati i solchi branchiali; segue una seconda cresta che nella porzione mediana, in corrispondenza della regione cardiaca, presenta un tratto quasi diritto più rilevato; la terza è incompleta e sviluppata solo ai lati; la quarta nella parte mediana ha forma di V molto aperta; la quinta cresta è incompleta ed è costituita da due tratti concavi ai lati della punta della V disegnata dalla quarta cresta.
Osservazioni. La presenza di un evidente solco circumgastrico permette di attribuire gli esemplari oggetto di studio ai Munidopsidae secondo la diagnosi del taxon fornita da Robinset al. (2013). Tra i generi compresi nella famiglia, le maggiori affinità sono quelle riscontrabili con i rappresentati di Faxegalathea Jakobsen & Collins, 1997 e Tethysmunida De Angeli & Ceccon, 2017, entrambi presenti nell’Eocene del Veneto occidentale, essenzialmente per le caratteristiche del rostro che è molto grande, spatolato, con margini spinosi, privo di carena mediana. Rispetto alle specie attibuite a Faxegalathea, la specie tipo F. platyspinosa Jakobsen & Collins, 1997 del Daniano della Danimarca e F. valeccensis Beschin, Busulini & Tessier, 2016 dello Ypresiano del Veneto, il nuovo taxon differisce per il numero di spine lungo i margini rostrali e soprattutto per la presenza delle creste trasversali continue presenti sulla parte posteriore del carapace. Le specie riferite al secondo genere, Tethysmunida corallina De Angeli & Ceccon, 2017 dello Ypresiano di Monte Magrè di Schio (Vicenza) e T. setifera Beschin, Busulini & Tessier, 2019 dell’Eocene superiore di Parona (Verona), rispetto al nuovo taxon presentano rostro proporzionalmente meno sviluppato, carapace più allargato con margini laterali più convessi e ornamentazione costituita da corte strie trasversali, da noduli e da porosità sparse ma non da creste trasversali continue (Beschin et al., 2016, 2019; De Angeli & Ceccon, 2017). Nella conformazione e ornamentazione del carapace si rilevano somiglianze anche con Spathagalathea minuta De Angeli & Garassino, 2002 dell’Eocene superiore del Veneto occidentale (De Angeli & Garassino, 2002; Beschin et al., 2019) la quale tuttavia è stata inserita nei Galatheidae per l’assenza del solco circumgastrico.
Le differenze su riportate giustificano l’istituzione di un nuovo genere di Munidopsidae.
Genere Faxegalathea Jakobsen & Collins, 1997
Specie tipo: Faxegalathea platyspinosa Jakobsen & Collins, 1997
Faxegalathea valeccensis Beschin, Busulini, Tessier, 2016
(fig. 21)
2016 Faxegalathea valeccensis Beschin, Busulini, Tessier in Beschin et al., p. 41, fig. 20, t. 4, ff. 2 a-c, 3.
2017 Faxegalathea valeccensis – Beschin et al., p. 10, t. 1, f. 1.
2017 Tethysmunida cf. T. valeccensis – De Angeli & Ceccon, p. 15, t. 3, ff.1-2.
2019 Faxegalathea valeccensis – Busulini et al., p. 20, figg. 1, 2.
Materiale e dimensioni. Sette carapaci: (CZ) MCZ 5898; (VdP) MCZ 5902 L: 2,0; MCZ 5901 L: 2,1; MCZ 5900 L: 2,9; MCZ 5899 L: 3,0; MCZ 5903; MCZ 5904. MCZ 5898 associato a Paratetralia convexa MCZ 6080; MCZ 5902 associato a Archaeotetra lessinea MCZ 6072; MCZ 5904 associato a Paratetralia convexa MCZ 6140.
Distribuzione. Eocene inferiore: Veneto (Altissimo, Monte Magrè di Schio – Vicenza; Bolca – Verona) (Beschin et al., 2016; De Angeli & Ceccon, 2017; Busulini et al., 2019).
Galatheoidea fam. indet., gen. indet., sp. indet. A
(fig. 22)
Materiale e dimensioni. Due propodi di chelipede: (CZ) MCZ 5915 h: 2,1 (sinistro); (VP) MCZ 5914 h: 3,2 (destro). MCZ 5915 con Eotrachinotocarcinus airaghii MCZ 5944.
Descrizione. Il palmo è rettangolare allungato, molto più lungo che alto. Margini superiore ed inferiore quasi diritti, appena divergenti; quello superiore porta quattro spine (e in MCZ 5915 anche un tubercolo distale), quello inferiore almeno sette. La superficie esterna, unica visibile, appare compressa e nel complesso liscia; porta a metà una serie longitudinale di quattro spine rivolte in avanti nella chela destra, una di cinque elementi in quella sinistra accompagnata da una seconda serie di tre nella metà inferiore alla base del dito fisso. Margine distale obliquo e concavo che continua col margine occlusale del dito fisso. Il dito fisso è conservato in MCZ 5915: è robusto, corto e triangolare.
Osservazioni. I resti ricordano Galatheidae indet. B di Parona (Beschin et al., 2019); possibili sono confronti con le chele degli esemplari attuali di Munida. Trattandosi di esemplari incompleti non è possibile procedere ad un inquadramento tassonomico definito.
Galatheoidea fam. indet., gen. indet., sp. indet. B
Materiale e dimensioni. Due frammenti di carapace: (VC) MCZ 5896 e (VdP) MCZ 5897.
Osservazioni. L’incompletezza dei resti con superficie parzialmente abrasa non consente una loro collocazione tassonomica più definita.
Famiglia Porcellanidae Haworth, 1825
Genere Disipia Beschin, Busulini & Tessier, 2016
Specie tipo: Disipia sorbinii Beschin, Busulini & Tessier, 2016
Disipia sorbinii Beschin, Busulini & Tessier, 2016
(fig. 23)
2016 Disipia sorbinii Beschin, Busulini & Tessier in Beschin et al., p. 42, fig. 21, t. 4, ff. 5a, b.
2019 Disipia sorbinii – Beschin et al., p. 80, fig. 22.
Materiale e dimensioni. Quattro carapaci: (VC) MCZ 5916 L: 2,5; MCZ 5917 L: 2,5; MCZ 6212 l: 3,2; MCZ 5918. MCZ 6212 associato a Paratetralia convexa MCZ 6097.
Osservazioni. De Angeli & Ceccon (2017) descrivono alcuni esemplari rinvenuti nello Ypresiano di Monte Magrè di Schio (Vicenza) che attribuiscono con incertezza alla specie soprattutto per alcune caratteristiche dei lobi epibranchiali.
Distribuzione. Eocene inferiore: Veneto (Rama di Bolca – Verona; Altissimo -Vicenza) (Beschin et al., 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
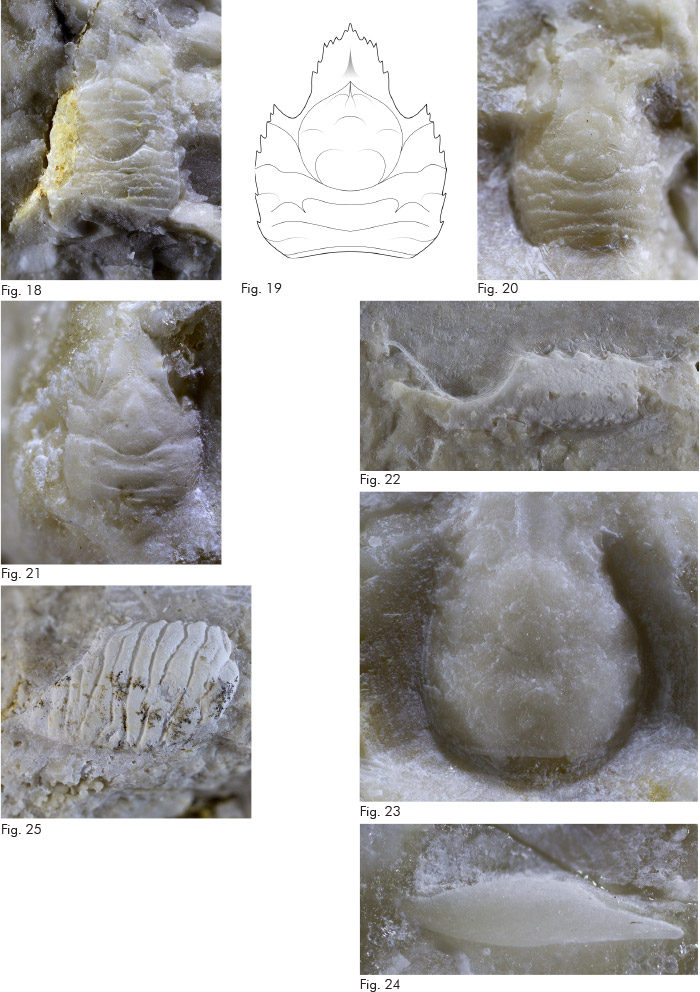
Fig. 19. Cristinagalathea striata gen. nov., sp. nov., profilo schematico di carapace.
Fig. 20. Cristinagalathea striata gen. nov., sp. nov., olotipo, es. MCZ 5906 (L: 2,6), carapace: visione dorsale.
Fig. 21. Faxegalathea valeccensis Beschin, Busulini, Tessier, 2016, es. MCZ 5901 (L: 2,1), carapace: visione dorsale.
Fig. 22. Galatheoidea fam. indet., gen. indet., sp. indet. A, es. MCZ 5915 (h: 2,1), propodo di chelipede sinistro: visione esterna.
Fig. 23. Disipia sorbinii Beschin, Busulini & Tessier, 2016, es. MCZ 5916 (L: 2,5), carapace: visione dorsale.
Fig. 24. Porcellanidae gen. indet., sp. indet., es. MCZ 5919 (lpr: 2,3 lp: 1,7), propodo di chelipede sinistro: visione interna.
Fig. 25. Dardanus braggensis Beschin, Busulini & Tessier, 2015, es. MCZ 5920 (lpr: 6,0 h: 6,0), propodo di chelipede sinistro: visione esterna.
Porcellanidae gen. indet., sp. indet.
(fig. 24)
Materiale e dimensioni. Un propodo di chelipede sinistro: (VC) MCZ 5919 lpr: 2,3 lp: 1,7 associato a Paguristes sp. MCZ 5923.
Descrizione. Propodo di chelipede sinistro di piccolissime dimensioni, allungato, con superficie convessa. Palmo più lungo che alto, subtriangolare e con margini superiore ed inferiore divergenti. Margine superiore concavo nel tratto rivolto al carpo e poi regolarmente convesso. Margine inferiore leggermente convesso. Margine distale obliquo, con ampia convessità presso la base del dito fisso. Dito fisso ben sviluppato, triangolare e con margine occlusale diritto. La ornamentazione della superficie non è rilevabile.
Osservazioni. Mancando la possibilità di osservare l’ornamentazione della superficie per le modestissime dimensioni dell’esemplare si preferisce non procedere a una più precisa determinazione ricordando però che propodi di aspetto simile sono già stati descritti per vari affioramenti dell’Eocene del Veneto (Beschin et al., 2016, 2018, 2019; De Angeli & Ceccon, 2017)
Superfamiglia Paguroidea Latreille, 1802
Famiglia Diogenidae Ortmann, 1892
Genere Dardanus Paul’son, 1875
Specie tipo: Dardanus hellerii Paul’son, 1875
Dardanus braggensis Beschin, Busulini & Tessier, 2015
(fig. 25)
2015 Dardanus braggensis Beschin, Busulini & Tessier, p. 54, t. 1, ff. 5 a, b.
2016 Dardanus braggensis – Beschin et al., p. 48, t. 5, f. 4.
Materiale e dimensioni. Un propodo di chelipede sinistro in visione esterna: (CZ) MCZ 5920 lpr: 6,0 h: 6,0.
Distribuzione. Eocene inferiore: Veneto (Vestenanova e Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2015, 2016).
Genere Paguristes Dana, 1851
Specie tipo: Paguristes hirtus Dana, 1851
Paguristes cfr. clampensis De Angeli & Caporiondo, 2017
(fig. 26)
2017 Paguristes clampensis De Angeli & Caporiondo, p. 15, fig. 8, t. 4, ff. 1-4.
Materiale e dimensioni. Un propodo di chelipede sinistro: (VP) MCZ 5922 lp: 3,0 h: 3,8.
Descrizione. Propodo di chelipede sinistro allungato, leggermente convergente. Palmo più alto che lungo, di sezione trasversale ellittica; articolazione carpo-propodiale subverticale, margine distale nettamente obliquo e concavo;
margine superiore poco convesso, di profilo acuto, con sei tubercoli appuntiti; margine inferiore quasi diritto. Dito fisso lungo e robusto, incompleto distalmente, ornato da tubercoli. Superficie esterna del palmo rigonfia; nella parte media e inferiore è ornata da numerosi tubercoli di varia dimensione non molto elevati, quella superiore è più scabrosa con almeno quattro tubercoli molto evidenti. La superficie interna, osservabile solo in fase di pulitura, mostrava palmo rigonfio, ornato nella parte distale da quattro tubercoli, e inserzione del dattilo ampia ed ovale seguita sul dito fisso da una concavità.
Osservazioni. La conformazione generale e l’ornamentazione permettono di avvicinare l’esemplare a Paguristes clampensis noto per l’Eocene medio di Cava Main di Arzignano (Vicenza); differisce però dalla serie tipo per il più alto numero di punte sul margine superiore del palmo e per la ornamentazione costituita da tubercoli più grossi e meno numerosi sulla parte superiore dello stesso e più abbondanti su quella inferiore.
Paguristes vallionensis Beschin, Busulini & Tessier, 2018
(figg. 27, 28)
2018 Paguristes vallionensis Beschin, Busulini & Tessier, p. 154, figg. 50, 51 a, b.
Materiale e dimensioni. Un propodo di chelipede destro in visione esterna: (VC) MCZ 5921 lp: 2,6 h: 2,6.
Diagnosi emendata. Propodo robusto con palmo subquadrato e rigonfio. Articolazione carpo-propodiale subverticale; margine superiore leggermente convesso con 6-7 spine; margine inferiore quasi diritto e liscio; margine distale molto inclinato. Dito fisso tozzo. Superficie esterna bombata, ricoperta di tubercoli; superficie interna con rigonfiamento trasversale e pochi tubercoli.
Emendeddiagnosis. Propodus stout with subsquared, swollen palm. Carpo-propodial articulation subvertical; upper margin weakly convex with 6-7 spines; lower margin almost straight, smooth; distal margin very slanting. Fixed finger stout. Outer surface swollen, covered with tubercles; inner surface with transverse swelling and few tubercles.
Descrizione. Propodo di chelipede piuttosto robusto e molto rigonfio con palmo subquadrato; margine prossimale subverticale, debolmente convesso, con parte dell’articolazione carpo-propodiale; margine superiore leggermente convesso con tubercolo prossimale e tracce di alcune spine; margine inferiore quasi diritto e liscio, poco convesso nel primo tratto poi concavo alla base del dito fisso; margine distale fortemente obliquo con una convessità a livello dell’articolazione con il dattilo. Il dito fisso è conservato solo in parte ma appare robusto e tozzo con margini debolmente convergenti. Tutta la superficie esterna del propodo è coperta da numerosi tubercoli inclinati in avanti distribuiti in modo irregolare, più evidenti presso il dito fisso.
Osservazioni. Il propodo rinvenuto ad Altissimo è più completo rispetto all’olotipo: la forma generale del palmo molto rigonfio e riccamente tubercolato con margine prossimale subverticale, quello superiore con alcune spine e quello distale fortemente obliquo, permettono di avvicinare però i due esemplari. Il dito fisso viene osservato per la prima volta e per questo si è deciso di fornire una descrizione emendata della specie che tiene conto anche di caratteri rilevati nell’olotipo proveniente da Campolongo di Val Liona (Vicenza) (Beschin et al., 2018).
Distribuzione. Eocene inferiore: Veneto (Altissimo – Vicenza); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza) (Beschin et al., 2018).
Paguristes sp.
(fig. 29)
Materiale e dimensioni. Un piccolo scudo: (VC) MCZ 5923 L: 3,5 l: 4,8; è associato a Porcellanidae indet. MCZ 5919.
Descrizione. Scudo allungato, convesso in senso trasversale, con massima larghezza nella parte anteriore. Margine fronto-orbitale conservato in piccola parte: si osserva solamente la cavità orbitale destra con una modesta punta extraorbitale. I margini laterali sono convessi nel primo tratto, poi quasi diritti e leggermente convergenti. Solco cervicale molto convesso. Cresta post-frontale molto convessa interessata da un solco longitudinale gastrico sottile e relativamente corto; solchi massetici molto incisi e un po’divergenti. Regione gastrica grande, di forma ovale, molto allungata, con la parte anteriore interessata dal citato solco longitudinale e coperta da brevi rugosità. Regioni massetiche allungate nel complesso triangolari.
Osservazioni. L’esemplare è incompleto a livello della fronte e nella parte posteriore dello scudo; mancando elementi indispensabili per la definizione specifica la sua attribuzione rimane indeterminata.
Famiglia Paguridae Latreille, 1802
Genere Anapagurus Henderson, 1886
Specie tipo: Pagurus laevis Bell, 1845
Anapagurus muelleri Beschin, Busulini & Tessier, 2018
(fig. 30)
1991 Anapagurus sp. – Müller & Collins, p. 54, fig. 2f, t. 1, f. 14.
2018 Anapagurus muelleri Beschin, Busulini & Tessier in Beschin et al., p. 156, ff. 52, 53.
2019 Anapagurus muelleri – Beschin et al., p. 89, fig. 42.
Materiale e dimensioni. Un propodo di chelipede sinistro (minore) in visione esterna: (VP) MCZ 5924 lpr: 4,5 lp: 3,1 h: 2,1.
Osservazioni. La specie è segnalata per la prima volta nell’Eocene inferiore.
Distribuzione. Eocene inferiore: Veneto (Altissimo – Vicenza); Eocene superiore: Ungheria (Müller & Collins, 1991) e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Famiglia Xylopaguridae Gašparič, Fraaije, Robin & De Angeli, 2016
Genere Lessinipagurus Beschin, De Angeli, Checchi & Zarantonello, 2012
Specie tipo: Lessinipagurus granulatus Beschin, De Angeli, Checchi & Zarantonello, 2012
Lessinipagurus cfr. bericus Beschin, Busulini & Tessier, 2018
(fig. 31)
2018 Lessinipagurus bericus Beschin, Busulini & Tessier in Beschin et al., p. 157, figg. 56, 57.
Materiale e dimensioni. Un propodo di chelipede destro in visione esterna: (VdP) MCZ 5925 lp: 3,6 h: 4,3.
Descrizione. Propodo di chelipede destro di modeste dimensioni, che ricorda nel complesso un crescente. Margini superiore ed inferiore convessi. Tutti i margini esterni del propodo appaiono spinosi ma quello superiore, meno convesso, presenta 6 o 7 spine più lunghe e acute. Angolo distale dorsale molto sviluppato e appuntito. Superficie quasi piatta piuttosto danneggiata; ornamentazione ricca costituita da numerosi nodi ovali particolarmente evidenti nella zona centrale accompagnati da altri più minuti rotondi; rilievo allungato e tubercolato all’attacco della superficie occlusale del dito fisso. Margine occlusale con un modesto dente distale.
Osservazioni. Le condizioni del propodo e le sue minute dimensioni, decisamente più piccole rispetto a quelle di tutti gli esemplari finora attribuiti a Lessinipagurus, non consentono un’attribuzione specifica sicura. I confronti effettuati con le varie specie del genere hanno mostrato le maggiori somiglianze con L. bericus conosciuto solo per l’Eocene superiore di Campolongo di Val Liona (Vicenza) (Beschin et al., 2018) unica specie che presenta delle spine lungo il margine superiore del propodo; l’olotipo, estremamente danneggiato, mostra però forma nel complesso più allungata e ornamentazione più regolare. L’esemplare di Altissimo è il più antico tra gli Xylopaguridae fossili che infatti vengono segnalati per la prima volta in terreni dell’Eocene inferiore.
Infraordine Brachyura Linnaeus, 1758
Sezione Dromiacea De Haan, 1833
Superfamiglia Homolodromioidea Alcock, 1900
Famiglia Goniodromitidae Beurlen, 1932
Genere Plagiophthalmus Bell, 1863
Specie tipo: Plagiophthalmus oviformis Bell, 1863
Plagiophthalmus paleogenicus (De Angeli & Ceccon, 2014) (comb. nov.)
(fig. 32)
2014 Dromiopsis paleogenicus De Angeli & Ceccon, 78, fig. 2(1-3).
Materiale e dimensioni. Un carapace incompleto: (VP) MCZ 5956 (L: 2,6) associato a Dromiopsis paucigranosa MCZ 5961.
Osservazioni. Le caratteristiche del carapace, anche se danneggiato nella regione frontale, consentono di attribuirlo con certezza alla specie. Come già evidenziato da Beschin et al. (2016) il taxon, originariamente attribuito al genere Dromiopsis (De Angeli & Ceccon, 2014), per la conformazione allungata del carapace, il margine frontale molto ampio e bifido, i solchi cervicale e branchiali ben distanziati e subparalleli, trova migliore collocazione in Plagiophthalmus.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza) (De Angeli & Ceccon, 2014).
Superfamiglia Dromioidea De Haan, 1833
Famiglia Dromiidae De Haan, 1833
Genere Dromiopsis Reuss, 1859
Specie tipo: Brachyurites rugosus Von Schlotheim, 1820
Dromiopsis ceratoi Beschin, Busulini & Tessier, 2016
(fig. 33)

Fig. 27. Paguristes vallionensis Beschin, Busulini & Tessier, 2018, profilo schematico di propodo.
Fig. 28. Paguristes vallionensis Beschin, Busulini & Tessier, 2018, MCZ 5921 (lp: 2,6 h: 2,6), propodo di chelipede destro: visione esterna.
Fig. 29. Paguristes sp., es. MCZ 5923 (L: 3,5 l: 4,8), scudo: visione dorsale.
Fig. 30. Anapagurus muelleri Beschin, Busulini & Tessier, 2018, es. MCZ 5924 (lpr: 4,5 lp: 3,1 h: 2,1), propodo di chelipede sinistro (minore): visione esterna.
Fig. 31. Lessinipagurus cfr. bericus Beschin, Busulini & Tessier, 2018, es. MCZ 5925 (lp: 3,6 h: 4,3), propodo di chelipede destro: visione esterna.
Fig. 32. Plagiophthalmus paleogenicus (De Angeli & Ceccon, 2014), es. MCZ 5956 (L: 2,6), carapace: visione dorsale.
Fig. 33. Dromiopsis ceratoi Beschin, Busulini & Tessier, 2016, es. MCZ 5950 (L: 5,7), carapace: visione dorsale.
2016 Dromiopsis ceratoi Beschin, Busulini & Tessier in Beschin et al., p. 59, fig. 32; t. 6, ff. 4, 5 a, b.
Materiale e dimensioni. Cinque carapaci: (CZ) MCZ 5951 L: 5,5; MCZ 5950 L: 5,7; MCZ 5949 L: 7,3; (VdP) MCZ 5952 L: 5,0; MCZ 5953.
Osservazioni. Negli esemplari decorticati i lobi epibranchiali sono più evidenti; si osserva inoltre la presenza di un dente bifido laterale a livello del solco cervicale che non era osservabile negli esemplari della serie tipo (Beschin et al., 2016).
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Dromiopsis longitudovata Beschin, Busulini & Tessier, 2016
(fig. 34)
2015 Dromiopsis sp. – Beschin et al., p. 60.
2016 Dromiopsis longitudovata Beschin, Busulini & Tessier in Beschin et al., p. 60, fig. 33; t. 6, ff. 6, 7 a, b.
Materiale e dimensioni. Due carapaci: (VP) MCZ 5955 L: 2,2 l: 2,4; MCZ 5954 L: 3,1 l: 3,2.
Distribuzione. Eocene inferiore: Veneto (Vestenanova, Rama di Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2015, 2016).
Dromiopsis marginospinosa Beschin, Busulini & Tessier, 2019
(fig. 35a, b)
2019 Dromiopsis marginospinosa Beschin, Busulini & Tessier in Beschin et al., p. 90, figg. 45, 46a, b.
Materiale e dimensioni. 12 carapaci: (CZ) MCZ 5931 L: 4,9 l: 3,8 Lo: 3,2; MCZ 5932 l: 7,4; MCZ 5933; (VC) MCZ 5938 L: 4,8 Lo: 2,9; MCZ 5936 L: 5,5 l: 4,0; MCZ 5937 L: 5,5; MCZ 5935 L: 9,5 l: 6,6 Lo: 4,9; MCZ 5934 L: 11,2; MCZ 5940 l: 2,9; MCZ 5939 l: 4,8; (VdP) MCZ 5942 L: 4,8 l: 3,8 Lo: 2,8; MCZ 5941 L: 11,6). MCZ 5938 associato a Branchioplax parva MCZ 6213.
Osservazioni. La specie è stata istituita sulla base di due minuscoli carapaci provenienti da Parona (Verona) (Beschin et al., 2019). Tra i numerosi esemplari rinvenuti ad Altissimo alcuni sono di dimensioni maggiori e permettono di osservare un aumento del rapporto L/l con la crescita. Per questo motivo gli esemplari più grandi ricordano Cyamocarcinus Bittner, 1883, ma se ne distinguono nettamente per la maggiore ampiezza delle orbite, che hanno margine sopraorbitale ondulato, e per la presenza dei solchi cervicale e branchiali che raggiungono i margini laterali. Si osserva inoltre il bordino finemente perlato sui margini antero-laterali, le orbite e la fronte, diagnostico della specie. Le caratteristiche di questo taxon, comunque consentono di avanzare ipotesi sulla stretta relazione filogenetica tra i generi Graptocarcinus Roemer, 1877, Dromiopsis e Cyamocarcinus.
Distribuzione. Eocene inferiore: Veneto (Altissimo – Vicenza); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Dromiopsis paucigranosa Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 36)
2007 Dromiopsis paucigranosa Beschin, Busulini, De Angeli & Tessier, p. 22, t. 2, ff. 1-3.
2011 Dromiopsis paucigranosa – Tessier et al., p. 215, f. 3.4.
2015 Dromiopsis paucigranosa – Beschin et al., p. 59, t. 2, f. 1.
2016 Dromiopsis paucigranosa – Beschin et al., p. 63, t. 7, f. 2.
2017 Dromiopsis paucigranosa – Beschin et al., p. 10, t. 1, f. 2.
Materiali e dimensioni. 13 carapaci: (VdP) MCZ 5957 L: 4,4; MCZ 5958; MCZ 5959; MCZ 5960; (VP) MCZ 5965 L: 5,0 l: 4,9; MCZ 5966 L: 5,0; MCZ 5964 L: 5,2; MCZ 5963 L: 5,3; MCZ 5962 L: 5,6; MCZ 5961 L: 5,7 l: 5,2; MCZ 5967; MCZ 5968; MCZ 5969. MCZ 5961 associato a Plagiophthalmus paleogenicus MCZ 5956.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011).
Dromiopsis sp.
Materiale e dimensioni. Un carapace: (VdP) MCZ 5970 associato a Cristinagalathea striata MCZ 5910 e MCZ 5911 e Prealpicarcinus cfr. laisensis MCZ 6015.
Osservazioni. L’esemplare risulta fortemente incompleto lungo i margini e mancante della fronte, ma la suddivisione in regioni del carapace ricorda quella di Dromiopsis paucigranosa.
Famiglia Dynomenidae Ortmann, 1892
Genere Kromtitis Müller, 1984
Specie tipo: Dromilites koberi Bachmayer & Tollmann, 1953
Kromtitis koberiformis Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 37)
2007 Kromtitis koberiformis Beschin, Busulini, De Angeli & Tessier, p. 26, t. 3, ff. 2-4.
2011 Kromtitis koberiformis – Tessier et al., p. 216.
2015 Kromtitis koberiformis – Beschin et al., p. 60.
2016 Kromtitis koberiformis – Beschin et al., p. 68, t. 8, f. 4.
2017 Kromtitis koberiformis – Beschin et al., p. 10, t. 1, f. 3.
Materiale e dimensioni. Un carapace incompleto (VP) MCZ 5971 (L > 10,3).
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011). Dubbia la sua presenza nell’Eocene superiore (Veneto, Campolongo di Val Liona – Vicenza) (Beschin et al., 2018)
Kromtitis pentagonalis Müller & Collins, 1991
(fig. 38)
1991 Kromtitis pentagonalis Müller & Collins, p. 63, t. 3, ff. 4, 8, non fig. 3e, t. 3, f. 5.
non 2016 Paradynomene pentagonalis – Beschin et al., p. 71, t. 8, f. 8.
2018 Kromtitis pentagonalis – Beschin et al., p. 160, fig. 61.
2019 Kromtitis pentagonalis – Beschin et al., p. 92, fig. 49.
Materiale e dimensioni. Tre carapaci: (VC) MCZ 5973 L: 4,9; MCZ 5972 L: 6,0; MCZ 5974.
Osservazioni. La specie viene segnalata per la prima volta in livelli dell’Eocene inferiore.
Distribuzione. Eocene inferiore: Veneto (Altissimo – Vicenza); Eocene superiore: Ungheria (Müller & Collins, 1991), Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Kromtitis subovatus Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 39)
2007 Kromtitis subovatus Beschin, Busulini, De Angeli & Tessier, p. 28, t. 3, ff. 6-8.
2016 Kromtitis subovatus – Beschin et al., p. 69, t. 8, f. 6 (cum syn.).
2018 Kromtitis cfr. subovatus – Beschin et al., p. 162, fig. 62.
2019 Kromtitis subovatus – Beschin et al., p. 93, fig. 50.
Materiale e dimensioni. Tre carapaci: (CZ) MCZ 5975 l: 5,5; MCZ 5977; (VC) MCZ 5976 L: 11,9.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Dromiacea incertae sedis sensu Schweitzer et al. (2012)
Genere Cyamocarcinus Bittner, 1883
Specie tipo: Cyamocarcinus angustifrons Bittner, 1883
Cyamocarcinus angustifrons Bittner, 1883
(fig. 40)

Fig. 35. Dromiopsis marginospinosa Beschin, Busulini & Tessier, 2019, es. MCZ 5942 (L: 4,8 l: 3,8 Lo: 2,8), carapace: a. visione dorsale, b. visione frontale.
Fig. 36. Dromiopsis paucigranosa Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 5961 (L: 5,7 l: 5,2), carapace: visione dorsale.
Fig. 37. Kromtitis koberiformis Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 5971 (misure parziali L: 10,3), carapace: visione dorsale.
Fig. 38. Kromtitis pentagonalis Müller & Collins, 1991, es. MCZ 5972 (L: 6,0), carapace: visione dorsale.
Fig. 39. Kromtitis subovatus Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 5976 (L: 11,9), carapace: visione dorsale.
Fig. 40. Cyamocarcinus angustifrons Bittner, 1883, es. MCZ 5929 (L: 10,5 Lo: 4,9), carapace: visione dorsale.
Fig. 41. Cyamocarcinus budensis Oppenheim, 1899, es. MCZ 5930 (L: 7,5 l: 5,5 Lo: 4,2), carapace: visione dorsale.
1883 Cyamocarcinus angustifrons Bittner, p. 310, t. 1, f. 8.
2006 Cyamocarcinus angustifrons – De Angeli & Garassino, p. 30 (cum syn.).
2016 Cyamocarcinus angustifrons – Beschin et al., p. 72, t. 9, f. 2 (cum syn.).
2017 Cyamocarcinus angustifrons – Beschin et al., p. 10, t. 1, f. 4, fig. 6.
2019 Cyamocarcinus angustifrons – Beschin et al., p. 93, fig. 52.
Materiale e dimensioni. Quattro carapaci: (CZ) MCZ 5826; (VC) MCZ 5927 L: 11,7 l: 7,4 Lo: 4,8; MCZ 5928; (VdP) MCZ 5929 L: 10,5 Lo: 4,9.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, contrada Gecchelina di Monte di Malo e San Pietro Mussolino – Vicenza; Bolca e Vestenanova – Verona) (Bittner, 1883; De Gregorio, 1895; Beschin et al., 2007,
2015, 2016; Tessier et al., 2011; Ceccon & De Angeli, 2013); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019), Sicilia (Checchia-Rispoli, 1905; Di Salvo, 1933) e Ungheria (Lőrenthey, 1898; Lőrenthey & Beurlen, 1929; Müller & Collins, 1991).
Cyamocarcinus budensis Oppenheim, 1899
(fig. 41)
1898 Cyamocarcinus angustifrons – Lőrenthey, p. 53, t. 3, f. 2.
1899 Cyamocarcinus budensis Oppenheim, p. 57.
2016 Cyamocarcinus budensis – Beschin et al., p. 73, t. 9, ff. 3 a, b (cum syn.).
Materiale e dimensioni. Un carapace: (VdP) MCZ 5930 L: 7,5 l: 5,5 Lo: 4,2.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); Eocene superiore: Ungheria (Lőrenthey, 1898; Lőrenthey & Beurlen, 1929).
Genere Eotrachynotocarcinus Beschin, Busulini, De Angeli & Tessier, 2007
Specie tipo: Eotrachynotocarcinus airaghii Beschin, Busulini, De Angeli & Tessier, 2007
Eotrachynotocarcinus airaghii Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 42)
2007 Eotrachynotocarcinus airaghii Beschin, Busulini, De Angeli & Tessier, p. 25, t. 2, ff. 7-9.
2011 Eotrachynotocarcinus airaghii – Tessier et al., p. 215, f. 3.5.
2015 Eotrachynotocarcinus airaghii – Beschin et al., p. 65, t. 2, f. 5.
2016 Eotrachynotocarcinus airaghii – Beschin et al., p. 74, 158, t. 9, ff. 4, 77.
2017 Eotrachynotocarcinus airaghii – Beschin et al., p. 10, t. 1, f. 5.
2019 Eotrachynotocarcinus airaghii – Pasini et al., p. 258, fig. 15B.
Materiale e dimensioni. Sei carapaci: (CZ) MCZ 5943; MCZ 5944; (VdP) MCZ 5946 L: 12,6 l: 10,0; MCZ 5945 L: 16,6; MCZ 5948 L > 4,7; MCZ 5947. MCZ 5944 in matrice con Galatheoidea indet.A MCZ 5915.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca -anche Pesciara- e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011).
Sezione Eubrachyura De Saint Laurent, 1980
Sottosezione Heterotremata Guinot, 1977
Superfamiglia Parthenopoidea MacLeay, 1838
Famiglia Parthenopidae MacLeay, 1838
Genere Mesolambrus Müller & Collins, 1991
Specie tipo: Mesolambrus declinatus Müller & Collins, 1991
Mesolambrus declinatus Müller & Collins, 1991
(fig. 43)
1991 Mesolambrus declinatus Müller & Collins, p. 69, fig. 3f, t. 3, ff. 11, 12, ?(14-17).
2016 Mesolambrus declinatus – Beschin et al., p. 83, t. 10, f. 4 (cum syn.).
2018 Mesolambrus declinatus – Beschin et al., p. 176, fig. 87.
2019 Mesolambrus declinatus – Beschin et al., p. 102, fig. 71.
Materiale e dimensioni. Tre carapaci: (VC) MCZ 5978; (VP) MCZ 5979 l: 6,8; MCZ 5980.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016); Eocene superiore: Ungheria e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Müller & Collins, 1991; Beschin et al., 2018, 2019).
Mesolambrus ypresianus Beschin, Busulini & Tessier, 2015
(fig. 44)
2015 Mesolambrus ypresianus Beschin, Busulini & Tessier, p. 69, fig. 7, t. 3, f. 5.
2016 Mesolambrus ypresianus – Beschin et al., p. 84, t. 10, f. 5.
Materiale e dimensioni. Un carapace incompleto: (VC) MCZ 5981 (L > 6,5).
Osservazioni. Il carapace, di dimensioni veramente modeste, è privo di buona parte della cuticola e questo non consente una puntuale osservazione della ricca tubercolazione di questa specie. Tuttavia si possono osservare, oltre alla forma
generale, la scarsa bombatura dei lobi protogastrici, le spine lungo i margini laterali e alcuni rigonfiamenti compatibili con la distribuzione dei tubercoli maggiori già rilevati in esemplari meglio conservati. L’attribuzione specifica risulta quindi sicura.
Distribuzione. Eocene inferiore: Veneto (Vestenanova e Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2015, 2016).
? Mesolambrus sp.
(fig. 45)
Materiale e dimensioni. Un carapace di piccole dimensioni: (VC) MCZ 5982 l: 2,3.
Descrizione. Carapace subesagonale, più largo che lungo, estremamente convesso nella parte anteriore. Margine frontale ampio, lamellare e quasi diritto ma con modesta incisione mediana e angolo esterno convesso. Orbite abbastanza grandi rivolte antero-lateralmente. Margine orbitale in continuità con quello frontale, intero, un po’ rigonfio; dente extraorbitale danneggiato. I margini antero-laterali sono mal conservati nel primo tratto ma nel complesso sono poco convessi e divergenti, ornati da alcuni denti spinosi: si possono osservare gli ultimi quattro che risultano piatti e molto sporgenti; l’ultimo, diretto all’esterno, si trova nel terzo posteriore del carapace. I margini postero-laterali sono più corti, concavi e decisamente convergenti. Il margine posteriore è quasi diritto, ma non del tutto conservato. La suddivisione in regioni è poco evidente per la cattiva conservazione della superficie ma si notano alcuni solchi poco profondi. Oltre le corte lamelle che costituiscono la fronte, i lobi protogastrici sono fortemente rigonfi e tra essi si intuisce il lobo mesogastrico con corpo relativamente stretto ma processo anteriore molto allungato, triangolare. I rami laterali del solco cervicale sono evidenti, decisamente convessi. La regione cardiaca è conservata solo in parte, abbastanza stretta e con margini indistinti. La suddivisione in lobi delle regioni branchiali risulta poco evidente: si nota però un solco branchiale obliquo che corre verso i margini postero-laterali.
Osservazioni. Le minute dimensioni del carapace e la cattiva conservazione della sua superficie, che non consente di osservare tracce di una eventuale tubercolazione, rendono molto difficile l’attribuzione dell’esemplare. La sua forma generale con fronte larga e lamellare, i margini antero-laterali spinosi e sporgenti che continuano fino al terzo posteriore, quelli postero-laterali decisamente concavi, così come le caratteristiche delle regioni centrali del dorso ricordano quelli dei rappresentanti di Mesolambrus, genere a cui viene dubitativamente attribuito. I lobi protogastrici estremamente bombati richiamano quelli di M. declinatus, specie comunque presente ad Altissimo di cui l’esemplare trattato potrebbe rappresentare una forma giovanile.
Superfamiglia Portunoidea Rafinesque, 1815
Famiglia Macropipidae Stephenson & Campbell, 1960
Genere Gecchelicarcinus Beschin, Busulini, De Angeli & Tessier, 2007
Specie tipo: Gecchelicarcinus lorigae Beschin, Busulini, De Angeli & Tessier, 2007
Gecchelicarcinus zanderigoi Beschin, Busulini & Tessier, 2016
(figg. 46, 47)
2016 Gecchelicarcinus zanderigoi Beschin, Busulini & Tessier in Beschin et al., p. 93, fig. 47; t. 11, ff. 8 a, b.
Materiale e dimensioni. Due carapaci: (CZ) MCZ 6010 L > 5,7; (VP) MCZ 6210 L: 7,1 l: 5,7.
Osservazioni. Anche se non ben conservati, i due esemplari si possono attribuire alla specie di cui era finora noto solo l’olotipo (Beschin et al., 2016). In particolare il carapace MCZ 6010 anche se incompleto consente di osservare in modo particolarmente evidente la ricca tubercolazione caratteristica del taxon.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Genere Vestenanovia Beschin, Busulini & Tessier, 2015
Specie tipo: Vestenanovia carinata Beschin, Busulini & Tessier, 2015
Vestenanovia carinata Beschin, Busulini & Tessier, 2015
(fig. 48)
2015 Vestenanovia carinata Beschin, Busulini & Tessier, p. 75, fig. 9, t. 4, ff. 4, 5.
2016 Vestenanovia carinata – Beschin et al., p. 95, t. 12, f. 1.
2018 Vestenanovia carinata – Beschin et al., p. 183, figg. 99, 100.
2019 Vestenanovia carinata – Beschin et al., p. 103, fig. 76.
Materiali e dimensioni. Quattro carapaci: (CZ) MCZ 5983 l: 6,0; MCZ 5984; (VC) MCZ 5986 l: 5,0; MCZ 5985 l: 11,7.
Distribuzione. Eocene inferiore: Veneto (Vestenanova e Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2015, 2016); Eocene superiore: Veneto (Parona – Verona e Campolongo di Val Liona – Vicenza) (Beschin et al., 2018, 2019).
Superfamiglia Carpilioidea Ortmann, 1893
Famiglia Carpiliidae Ortmann, 1893
Genere Braggicarpilius Beschin, Busulini & Tessier, 2015
Specie tipo: Braggicarpilius marginatus Beschin, Busulini & Tessier, 2015
Braggicarpilius marginatus Beschin, Busulini & Tessier, 2015
(fig. 49)
2015 Braggicarpilius marginatus Beschin, Busulini & Tessier, p. 79, fig. 11, t. 5, f. 1.
2016 Braggicarpilius marginatus – Beschin et al., p. 97, t. 12, f. 4.
2018 Braggicarpilius marginatus – Beschin et al., p. 183, figg. 102 a, b.
Materiale e dimensioni. Un carapace in parte corroso: (VdP) MCZ 6211 L: 4,3 l: 3,4.
Osservazioni. Il carapace è corroso sulla superficie dorsale e lungo i margini; la fronte, inoltre, non è chiaramente individuata. Tuttavia la forma generale dell’esemplare consente di attribuirlo a questa specie. Le orbite si presentano infatti ampie e ben evidenti sono anche i solchi branchio-cardiaci.
Distribuzione. Eocene inferiore: Veneto (Vestenanova e Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2015, 2016); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza) (Beschin et al., 2018).
Genere Montemagralia De Angeli & Ceccon, 2016
Specie tipo: Montemagralia lata De Angeli & Ceccon, 2016
Osservazioni. Il taxon, originariamente inserito tra i Trapeziidae (De Angeli & Ceccon, 2016a), è stato attribuito ai Carpiliidae da Schweitzer et al. (2018) nonostante abbia la fronte diritta, scelta che qui viene seguita vista la forma generale del carapace. Riteniamo comunque che la posizione sistematica del genere debba essere ulteriormente indagata.
? Montemagralia cfr. lata De Angeli & Ceccon, 2016
(fig. 50)
carapace: visione dorsale.
Fig. 43. Mesolambrus declinatus Müller & Collins, 1991, es. MCZ 5979 (l: 6,8), carapace: visione dorsale.
Fig. 44. Mesolambrus ypresianus Beschin, Busulini & Tessier, 2015, es. MCZ 5981 (misura parziale L: 6,5), carapace: visione dorsale.
Fig. 45. ? Mesolambrus sp., es. MCZ 5982 (l: 2,3), carapace: visione dorsale.
Fig. 46. Gecchelicarcinus zanderigoi Beschin, Busulini & Tessier 2016, es. MCZ 6210 (L: 7,1 l: 5,7), carapace: visione dorsale.
Fig. 47. Gecchelicarcinus zanderigoi Beschin, Busulini & Tessier 2016, es. MCZ 6010 (misura parziale L: 5,7), carapace: visione dorsale.
Fig. 48. Vestenanovia carinata Beschin, Busulini & Tessier, 2015, es. MCZ 5983 (l: 6,0), carapace: visione dorsale.
Fig. 49. Braggicarpilius marginatus Beschin, Busulini & Tessier, 2015, es. MCZ 6211 (L: 4,3 l: 3,4), carapace: visione dorsale.
2016a Montemagralia lata De Angeli & Ceccon, p. 130, fig. 2(1 – 3).
2018 Montemagralia lata – Schweitzer et al., p. 5, fig. 4(5b).
Materiale e dimensioni. Una porzione di carapace: (CZ) MCZ 5987 (L > 3,9).
Osservazioni. Il resto è estremamente parziale e conserva solamente la parte destra del carapace con l’orbita e il margine laterale. Profilo del carapace simile si riscontra in più specie rinvenute in livelli ypresiani del Veneto come Corallicarpilus arcuatus De Angeli & Ceccon, 2015 e Montemagralia lata, entrambe note per Monte Magrè di Schio (Vicenza). L’orbita relativamente grande rende il reperto analizzato più vicino alla seconda cui viene attribuito comunque con un margine di incertezza.
Genere Paraocalina Beschin, Busulini, De Angeli & Tessier, 2007
Specie tipo: Paraocalina multilobata Beschin, Busulini, De Angeli & Tessier, 2007
Osservazioni. Il genere viene inserito tra i Carpiliidae secondo quanto proposto nel lavoro istitutivo e nel recente Treatise Online (Beschin et al., 2007; Schweitzer et al., 2018). De Angeli & Ceccon (2016b) hanno invece accostato il taxon agli Xanthidae sulla base di esemplari che non sembrano coincidere con quelli della serie tipo di P. multilobata.
Paraocalina cfr. silviae Beschin, Busulini, De Angeli & Tessier, 2016
(fig. 51)
2016 Paraocalina silviae Beschin, Busulini, De Angeli & Tessier in Beschin et al., p. 98, fig. 49, t. 12, ff. 7, 8.
Materiale e dimensioni. Un frammento di carapace: (CC) MCZ 6017 L > 1,9.
Osservazioni. Nonostante del carapace sia conservata solamente la parte laterale sinistra e una porzione di quella centrale, l’identificazione appare abbastanza sicura per la presenza della denticolazione sul margine antero-laterale, di granuli sulla superficie e di una leggera cresta che parte dall’angolo laterale.
Distribuzione. Eocene inferiore: Veneto (Bolca e probabilmente Altissimo – Vicenza) (Beschin et al., 2016)
Famiglia Palaeoxanthopsidae Schweitzer, 2003
Genere Latuxanthides De Angeli & Ceccon, 2015
Specie tipo: Latuxanthides dentatus De Angeli & Ceccon, 2015
Latuxanthides dentatus De Angeli & Ceccon, 2015
(fig. 52)
2015 Latuxanthides dentatus De Angeli & Ceccon, p. 127, fig. 5(1, 2).
Materiale e dimensioni. Un carapace: (VC) MCZ 5988 l: 6,7.
Osservazioni. Sulla base della conformazione delle regioni posteriori del carapace, De Angeli & Ceccon (2015) attribuiscono alla specie, se pur in modo dubitativo, anche ? Lobulata sp. dello Ypresiano di contrada Gecchelina di Monte di Malo (Vicenza) di Beschin et al. (2007). Tuttavia l’esemplare citato appare nel complesso meno allargato, con margini postero-laterali un po’ convessi anziché concavi e dente laterale triangolare poco sporgente, per cui l’ipotesi non appare condivisibile.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza) (De Angeli & Ceccon, 2015).
Famiglia Tumidocarcinidae Schweitzer, 2005
Genere Titanocarcinus A. Milne Edwards, 1863
Specie tipo: Titanocarcinus serratifrons A. Milne Edwards, 1863
Titanocarcinus raulinianus A. Milne Edwards, 1863
(fig. 53)
1863 Titanocarcinus raulinianus A. Milne Edwards, p. 277, t. 17, ff. 3, 4.
1929 Titanocarcinus raulinianus – Lőrenthey & Beurlen, p. 234, t. 11, ff. 6, 7.
1933 Titanocarcinus sp. – Di Salvo, p. 30, t. 2, f. 9. 2006 Titanocarcinus raulinianus – De Angeli & Garassino, p. 73 (cum syn.).
2007 Titanocarcinus raulinianus – Beschin et al., p. 52, t. 7, ff. 4-6.
2016 Titanocarcinus raulinianus – Beschin et al., p. 104, t. 13, f. 3.
2018 Titanocarcinus raulinianus – Beschin et al., p. 186, fig. 106.
Materiale e dimensioni. Tre carapaci incompleti: (CC) MCZ 5989; (VC) MCZ 5990 l: 4,3; (VdP) MCZ 5991. MCZ 5990 associato a Cristinagalathea striata MCZ 5905.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca – Verona) (Beschin et al., 2007, 2016); Eocene medio: Francia (Milne Edwards, 1863), Puglia, Sicilia e Friuli (Di Salvo, 1933; De Angeli & Garassino, 2006); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza) (Beschin et al., 2018), Ungheria (Lőrenthey & Beurlen, 1929).
Superfamiglia Pilumnoidea Samouelle, 1819
Famiglia Pilumnidae Samouelle, 1819
Genere Galenopsis A. Milne Edwards, 1865
Specie tipo: Galenopsis typica A. Milne Edwards, 1865
Galenopsis similis Bittner, 1875
(fig. 54)
1875 Galenopsis similis Bittner, p. 97, t. 2, f. 9.
2006 Galenopsis similis – De Angeli & Garassino, p. 71 (cum syn.).
2016 Galenopsis similis – Beschin et al., p. 107, t. 13, f. 6 (cum syn.).
2018 Galenopsis similis – Beschin et al., p. 188, fig. 109.
2019 Galenopsis similis – Beschin et al., p. 106, fig. 81.
Materiale e dimensioni. Cinque carapaci: (VdP) MCZ 5997 L: 6,8 l: 5,2; (VP) MCZ 5994 L: 33,0; MCZ 5993 L: 37,5; MCZ 5996 l: 22,2; MCZ 5995 l: 26,1. Gli esemplari MCZ 5994 e MCZ 5996 sono conservati nella stessa matrice.
Osservazioni. L’esemplare MCZ 5997 conserva anche la chela sinistra in connessione anatomica e resti di altri pereiopodi: le caratteristiche del chelipede corrispondono a quanto già osservato da Beschin et al. (2018).
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Monte Magrè di Schio, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Bittner, 1875; Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); Eocene superiore: Ungheria (Müller & Collins, 1991), Sicilia (Di Salvo, 1933) e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Genere Glabropilumnus Balss, 1932
Specie tipo: Xantho dispar Dana, 1852
Glabropilumnus trispinosus Beschin, Busulini & Tessier, 2016
(fig. 55)
2016 Glabropilumnus trispinosus Beschin, Busulini & Tessier in Beschin et al., p. 108 , fig. 52; t. 14, f. 1
Materiale e dimensioni. Un carapace: (VdP) MCZ 5998 L: 4,2 l: 3,5.
Osservazioni. Come nell’olotipo, rinvenuto nello Ypresiano di Bolca (Beschin et al., 2016), l’esemplare conserva un peduncolo oculare.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Genere Lobogalenopsis Müller & Collins, 1991
Specie tipo: Galenopsis quadrilobatus Lőrenthey, 1898
Lobogalenopsis quadrilobata (Lőrenthey, 1898)
(fig. 56)
1897 Galenopsis quadrilobatus Lőrenthey, p. 156 (nom. nud.).
1898 Galenopsis quadrilobatus Lőrenthey, p. 66, t. 5, f. 3.
1991 Lobogalenopsis quadrilobata – Müller & Collins, p. 88, fig. 5f, t. 8, ff. 8-10.
2006 Lobogalenopsis quadrilobata – De Angeli & Garassino, p. 71 (cum syn.).
2016 Lobogalenopsis quadrilobata – Beschin et al., p. 109, t. 13, ff. 7, 8 (cum syn.).
2018 Lobogalenopsis quadrilobata – Beschin et al., p. 188, fig. 110.
2019 Lobogalenopsis quadrilobata – Beschin et al., p. 107, fig. 85.
Materiale e dimensioni. Nove carapaci: (CC) MCZ 6216 L: 8,2; MCZ 5999; (CZ) MCZ 6000; (VdP) MCZ 6001 L: 7,3; MCZ 6002; (VP) MCZ 6003 L: 6,5 l: 5,3; MCZ 6004 l: 8,9; MCZ 6005; MCZ 6006.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino e Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); Eocene superiore: Ungheria (Lőrenthey, 1897,1898; Müller & Collins, 1991), Sicilia (Di Salvo, 1933) e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Genere Palladiocarcinus De Angeli & Ceccon, 2014
Specie tipo: Palladiocarcinus brevidentatus De Angeli & Ceccon, 2014
Palladiocarcinus brevidentatus De Angeli & Ceccon, 2014
(fig. 57)
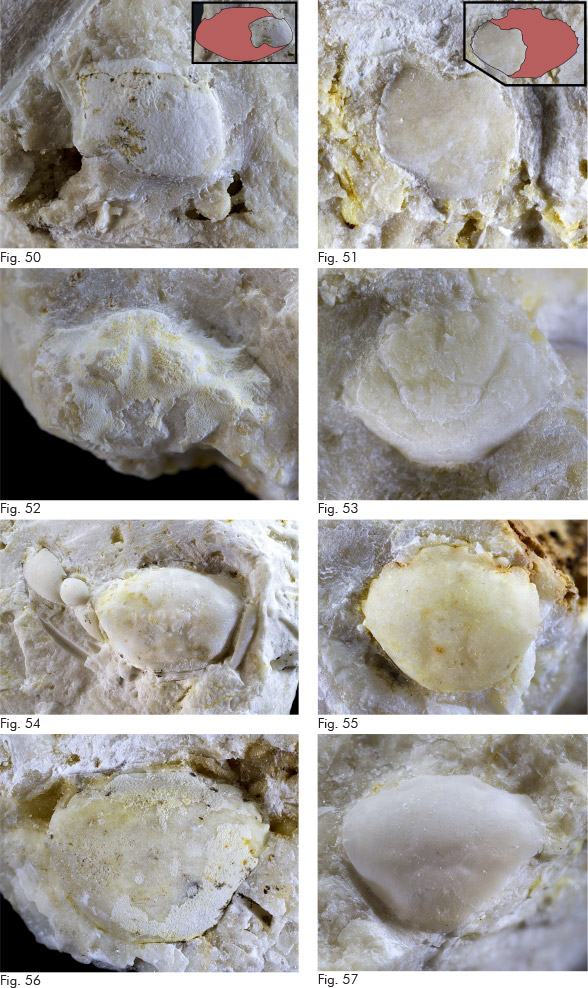
Fig. 51. Paraocalina cfr. silviae Beschin, Busulini, De Angeli & Tessier, 2016, es. MCZ 6017 (misura parziale L: 1,9), frammento di carapace.
Fig. 52. Latuxanthides dentatus De Angeli & Ceccon, 2015, es. MCZ 5988 (l: 6,7), carapace: visione dorsale.
Fig. 53. Titanocarcinus raulinianus A. Milne Edwards, 1863, es. MCZ 5990 (l: 4,3), carapace: visione dorsale.
Fig. 54. Galenopsis similis Bittner, 1875, es. MCZ 5997 (L: 6,8 l: 5,2), carapace in visione dorsale e chelipede sinistro.
Fig. 55. Glabropilumnus trispinosus Beschin, Busulini & Tessier, 2016, es. MCZ 5998 (L: 4,2 l: 3,5), carapace: visione dorsale.
Fig. 56. Lobogalenopsis quadrilobata (Lőrenthey, 1898), es. MCZ 6003 (L: 6,5 l: 5,3), carapace: visione dorsale.
Fig. 57. Palladiocarcinus brevidentatus De Angeli & Ceccon, 2014, es. MCZ 6007 (L: 9,6 l: 7,5), carapace: visione dorsale.
2014 Palladiocarcinus brevidentatus De Angeli & Ceccon, p. 87, ff. 5(1-3).
2015 Palladiocarcinus brevidentatus – Beschin et al., p. 85, t. 6, f. 2.
2016 Palladiocarcinus brevidentatus – Beschin et al., p. 110, t. 14, f. 2.
Materiale e dimensioni. Un carapace: (VC) MCZ 6007 L: 9,6 l: 7,5.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (De Angeli & Ceccon, 2014; Beschin et al., 2015, 2016).
Genere Prealpicarcinus De Angeli & Ceccon, 2015
Specie tipo: Prealpicarcinus dallagoi De Angeli & Ceccon, 2015
Prealpicarcinus cfr. laisensis Beschin, Busulini & Tessier, 2016
(fig. 58)
2016 Prealpicarcinus laisensis Beschin, Busulini & Tessier in Beschin et al., p. 110, fig. 53, t. 14, ff. 3 a, b.
Materiale e dimensioni. Un carapace di dimensioni molto piccole: (VdP) MCZ 6015 L: 1,9; è associato a Cristinagalathea striata MCZ 5910 e MCZ 5911 e Dromiopsis sp. MCZ 5970.
Osservazioni. Il minuscolo carapace, incompleto posteriormente e cristallizzato, mostra una suddivisione delle regioni simile a quelle osservabili negli esemplari della serie tipo, proveniente da accumuli organogeni ypresiani dell’area di Bolca (Vicenza e Verona) (Beschin et al., 2016), in modo particolare in quelli conservati in modo analogo al reperto analizzato che è completamente decorticato. L’incertezza nell’attribuzione è dovuta alla incompletezza del carapace.
Superfamiglia Trapezioidea Miers, 1886
Famiglia Tetraliidae Castro, Ng & Ahyong, 2004
Genere Scutata Beschin, Busulini & Tessier in Beschin et al., 2016
Specie tipo: Scutata eocenica Beschin, Busulini & Tessier, 2016
Scutata eocenica Beschin, Busulini & Tessier, 2016
(fig. 59)
2016 Scutata eocenica Beschin, Busulini & Tessier in Beschin et al., p. 123, fig. 60, t. 15, ff . 9a, b.
Materiale e dimensioni. Cinque carapaci: (VdP) MCZ 6050 L: 5,5 l: 4,7; MCZ 6049 L: 9,2; MCZ 6051 l: 4,5; (VP) MCZ 6053 L: 4,0 l: 3,1; MCZ 6052 L: 5,7. MCZ 6050 associato a Vicetitrapezia exaltissimo MCZ 6202.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Famiglia Trapeziidae Miers, 1886
Genere Archaeotetra Schweitzer, 2005
Specie tipo: Archaeotetra inornata Schweitzer, 2005
Archaeotetra lessinea De Angeli & Ceccon, 2013
(fig. 60)
2013b Archaeotetra lessinea De Angeli & Ceccon, p. 31, f. 4 (1-5).
2016 Archaeotetra lessinea – Beschin et al., p. 125, t. 16, ff. 3 a, b.
Materiale e dimensioni. 20 carapaci: (CC) MCZ 6054 – MCZ 6056; (CZ) MCZ 6058 L: 8,8; MCZ 6057 L: 9,4 l: 6,4; MCZ 6059 l: 5,0; MCZ 6060; MCZ 6061; (VC) MCZ 6063 L: 5,2; MCZ 6062 L: 9,3; MCZ 6064 l: 4,5; MCZ 6065; MCZ 6066; (VdP) MCZ 6069 L: 4,6; MCZ 6067 L: 6,9 l: 5,1; MCZ 6068 L: 6,9; MCZ 6070 l: 4,5; MCZ 6071; MCZ 6072; (VP) MCZ 6073 L: 5,4. MCZ 6064 associato a Branchioplax cordata MCZ 6209; MCZ 6072 a Faxegalathea valeccensis MCZ 5902.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza; Bolca – Verona) (De Angeli & Ceccon, 2013b; Beschin et al., 2016).
Genere Eomaldivia Müller & Collins, 1991
Specie tipo: Eomaldivia trispinosa Müller & Collins, 1991
Eomaldivia trispinosa Müller & Collins, 1991
(fig. 61)
1991 Eomaldivia trispinosa Müller & Collins, p. 81, fig. 4l, t. 6, ff. 8, 11.
2013b Eomaldivia trispinosa – De Angeli & Ceccon, p. 33, fig. 5(1-4).
2018 Eomaldivia trispinosa – Beschin et al., p. 192, fig. 119.
2019 Eomaldivia trispinosa – Beschin et al., p. 111, fig. 95.
Materiale e dimensioni. Tre carapaci: (VC) MCZ 6074 L: 4,4 l: 3,5; (VdP) MCZ 6075 L > 6,8; (VP) MCZ 6076 L: 7,0.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza) (De Angeli & Ceccon, 2013b); Eocene superiore: Ungheria (Müller & Collins, 1991) e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Genere Paratetralia Beschin, Busulini, De Angeli & Tessier, 2007
Specie tipo: Paratetralia convexa Beschin, Busulini, De Angeli & Tessier, 2007
Paratetralia convexa Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 62)
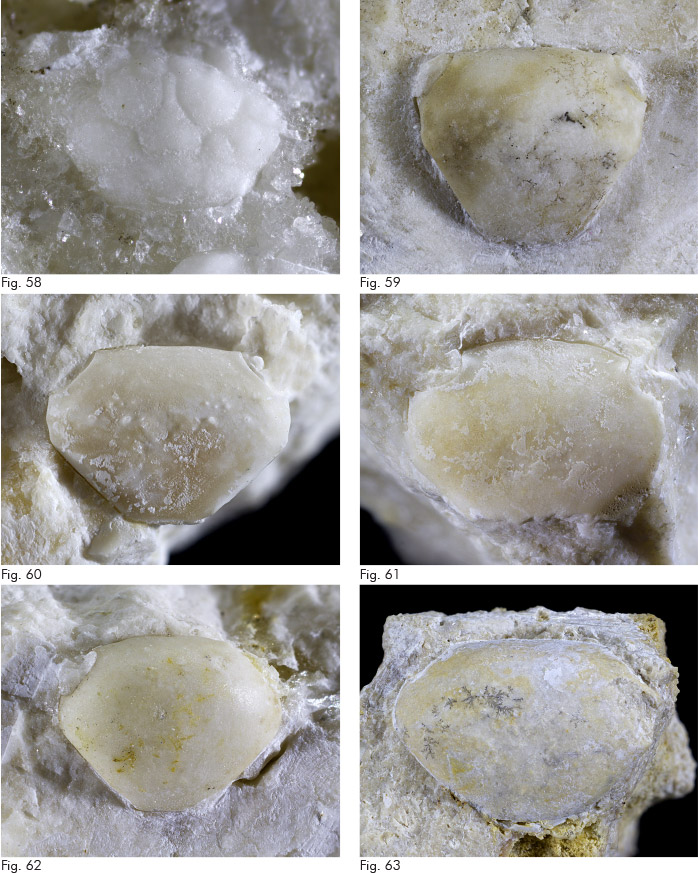
Fig. 59. Scutata eocenica Beschin, Busulini & Tessier, 2016, es. MCZ 6050 (L: 5,5 l: 4,7), carapace: visione dorsale.
Fig. 60. Archaeotetra lessinea De Angeli & Ceccon, 2013, es. MCZ 6067 (L: 6,9 l: 5,1), carapace: visione dorsale.
Fig. 61. Eomaldivia trispinosa Müller & Collins, 1991, es. MCZ 6075 (misura parziale L: 6,8), carapace: visione dorsale.
Fig. 62. Paratetralia convexa Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6117 (L: 4,6 l: 3,7), carapace: visione dorsale.
Fig. 63. Paratetralia sulcata De Angeli & Ceccon, 2013, es. MCZ 6182 (l: 8,9), carapace: visione dorsale.
2007 Paratetralia convexa Beschin, Busulini, De Angeli & Tessier, p. 55, t. 8, ff. 6a-c, 7, 8a, b.
2011 Paratetralia convexa – Tessier et al., p. 218, f. 3.9a.b.
2013b Paratetralia convexa – De Angeli & Ceccon, p. 34, f. 6 (1-4).
2015 Paratetralia convexa – Beschin et al., p. 87, t. 6, f. 5.
2016 Paratetralia convexa – Beschin et al., p. 125, t . 16, f. 4,
Materiale e dimensioni. 105 carapaci: (CC) MCZ 6078 L: 4,3 l: 3,6; MCZ 6077 L: 12,0 l: 5,0; MCZ 6079; (CZ) MCZ 6084 L: 3,0 l: 2,6; MCZ 6083 L: 4,2; MCZ 6082 L: 7,3; MCZ 6081 L: 7,4 l: 5,2; MCZ 6080 L: 7,6; MCZ 6085; MCZ 6086; (VC) MCZ 6088 L: 4,4; MCZ 6087 L: 5,5; MCZ 6103 l: 3,0; MCZ 6102 l: 3,1; MCZ 6101 l: 3,2; MCZ 6100 l: 3,5; MCZ 6099 l: 4,1; MCZ 6097 l: 4,4; MCZ 6098 l: 4,4; MCZ 6096 l: 4,5; MCZ 6095 l: 4,6; MCZ 6094 l: 5,3; MCZ 6093 l: 5,6; MCZ 6092 l: 5,7; MCZ 6091 l: 6,0; MCZ 6090 l: 6,4; MCZ 6089 l: 6,7; MCZ 6104 – MCZ 6108; (VdP) MCZ 6121 L: 4,0 l: 3,3; MCZ 6120 L: 4,3; MCZ 6119 L: 4,4; MCZ 6118 L: 4,5; MCZ 6117 L: 4,6 l: 3,7; MCZ 6116 L: 4,7; MCZ 6115 L: 4,8 l: 3,8; MCZ 6114 L: 4,9; MCZ 6113 L: 5,2; MCZ 6112 L: 6,5 l: 5,0; MCZ 6111 L: 7,4; MCZ 6110 L: 8,5 l: 5,9; MCZ 6109 L: 8,9; MCZ 6130 l: 2,1; MCZ 6129 l: 2,2; MCZ 6128 l: 2,8; MZC 6127 l: 4,1; MCZ 6126 l: 4,2; MCZ 6125 l: 4,8; MCZ 6124 l: 5,3; MCZ 6123 l: 5,6; MCZ 6122 l: 6,2; MCZ 6131 – MCZ 6140; (VP) MCZ 6158 L: 3,8; MCZ 6157 L: 3,9; MCZ 6156 L: 4,5; MCZ 6155 L: 4,8; MCZ 6154 L: 4,9; MCZ 6153 L: 5,0 l: 4,2; MCZ 6152 L: 5,0 l: 4,5; MCZ 6151 L: 5,2 l: 4,1; MCZ 6150 L: 5,9; MCZ 6149 L: 6,1; MCZ 6148 L: 6,1 l: 4,6; MCZ 6147 L: 6,7 l: 5,0; MCZ 6146 L: 6,9; MCZ 6145 L: 7,0; MCZ 6144 L: 8,0; MCZ 6143 L: 8,1; MCZ 6142 L: 8,2; MCZ 6141 L: 9,3 l: 6,9; MCZ 6170 l: 2,7; MCZ 6169 l: 2,9; MCZ 6168 l: 3,4; MCZ 6167 l: 4,1; MCZ 6166 l: 4,3; MCZ 6165 l: 4,8; MCZ 6164 l: 5,0; MCZ 6162 l: 5,1; MCZ 6163 l: 5,1; MCZ 6161 l: 5,7; MCZ 6160 l: 6,2; MCZ 6159 l: 7,4; MCZ 6171 – MCZ 6181.
MCZ 6080 associato a Faxegalathea valeccensis MCZ 5898; MCZ 6092 a Prochlorodius ellipticus MCZ 6044; MCZ 6097 a Disipia sorbinii MCZ 6212; MCZ 6102 a Dromiopsis marginospinosa MCZ 5939; MCZ 6114 a Cristinagalathea striata MCZ 5907; MCZ 6140 a Faxegalathea valeccensis MCZ 5904. MCZ 6154 associato a MCZ 6163; MCZ 6165 associato a MCZ 6168.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Monte Magrè di Schio, Altissimo – Vicenza; Vestenanova, Bolca – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011; De Angeli & Ceccon, 2013b).
Paratetralia sulcata De Angeli & Ceccon, 2013
(fig. 63)
Fig. 59. Scutata eocenica Beschin, Busulini & Tessier, 2016, es. MCZ 6050 (L: 5,5 l: 4,7), carapace: visione dorsale.
Fig. 60. Archaeotetra lessinea De Angeli & Ceccon, 2013, es. MCZ 6067 (L: 6,9 l: 5,1), carapace: visione dorsale.
Fig. 61. Eomaldivia trispinosa Müller & Collins, 1991, es. MCZ 6075 (misura parziale L: 6,8), carapace: visione dorsale.
Fig. 62. Paratetralia convexa Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6117 (L: 4,6 l: 3,7), carapace: visione dorsale.
Fig. 63. Paratetralia sulcata De Angeli & Ceccon, 2013, es. MCZ 6182 (l: 8,9), carapace: visione dorsale.
2013b Paratetralia sulcata De Angeli & Ceccon, p. 36, f. 7 (1, 2).
2016 Paratetralia sulcata – Beschin et al., p. 126, t. 16, f. 5.
Materiale e dimensioni. 12 carapaci: (CC) MCZ 6182 l: 8,9; (CZ) MCZ 6185 L: 5,5; MCZ 6184 L: 8,5; MCZ 6183 L: 10,3; MCZ 6186; MCZ 6187; (VC) MCZ 6190 L: 8,5; MCZ 6189 L: 8,6 l: 6,0; MCZ 6188 L: 10,9; (VdP) MCZ 6191 l: 9,5; (VP) MCZ 6193 L: 6,7 l: 5,3; MCZ 6192 L: 11,9.
Distribuzione. Eocene inferiore: Veneto (Monte Magrè di Schio, Altissimo – Vicenza; Bolca – Verona) (De Angeli & Ceccon, 2013b; Beschin et al., 2016).
Famiglia Vicetitrapeziidae fam. nov.
Genere tipo: Vicetitrapezia gen. nov.
Diagnosi. Carapace liscio, molto convesso in senso longitudinale. Fronte molto ampia, rivolta verso il basso, con margine bilobo, debolmente ondulato. Orbite rivolte antero-lateralmente; dente extraorbitale robusto. Margini antero-laterali terminanti con concavità semicircolare; margini postero-laterali nettamente convergenti; margine posteriore diritto e più stretto della fronte. Regioni dorsali non definite.
Diagnosis. Carapace smooth, longitudinally strongly convex. Front very broad, downward folded, with bilobed, weakly wavy margin. Orbits anterolateral directed; outer orbital tooth stout. Anterolateral margins with semicircular concavity at the end; posterolateral ones straight and strongly convergent; posterior margin straight and narrower than front. Dorsal regions not defined.
Osservazioni. Castro et al. (2004), sulla base di considerazioni che coinvolgono anche caratteri che non si conservano allo stato fossile come per esempio l’addome degli individui di sesso maschile, hanno suddiviso i Trapeziidae sensu Miers, 1886, piccoli brachiuri adattati a vivere in ambiente corallino, in tre famiglie e precisamente Domeciidae Ortmann, 1893, Tetraliidae e Trapeziidae. Considerando quindi solamente i caratteri relativi alla conformazione del carapace, i domecidi presentano carapace ovale allargato, margine frontale quasi rettilineo, liscio o finemente denticolato, margini antero-laterali con alcuni denti o piccoli tubercoli, margine posteriore largo all’incirca come la fronte, regioni del dorso non distinte e superficie liscia o granulata; in terreni paleogenici del Veneto la famiglia è rappresentata da Cherusius oligocenicus (Beschin, De Angeli & Checchi, 2001), Palmyria levigata Beschin, Busulini & Tessier, 2016 e Proticalia parva (Beschin, Busulini & Tessier, 2016)(De Angeli & Garassino, 2006; Beschin et al., 2016; De Angeli & Ceccon, 2017; De Angeli & Caporiondo, 2020). I tetralidi presentano invece carapace ovale allungato, relativamente appiattito, il margine frontale quasi diritto, in genere finemente denticolato, spine antero-laterali assenti negli individui adulti, margini postero-laterali fortemente convergenti, superficie del dorso liscia e regioni non distinte; nel Paleogene del Veneto sono presenti: Eurotetralia loerenthey (Müller, 1975), Scutata eocenica Beschin, Busulini & Tessier, 2016, Tetralia minuta Beschin, Busulini & Tessier, 2016, Tetralia vicetina De Angeli & Ceccon, 2013 (De Angeli et al., 2019). Nei trapezidi il carapace può avere forma varia da esagonale ad ovale allargata; la fronte si presenta in genere lobata o con larghi denti, i margini antero-laterali sono lisci o con una spina, margine posteriore è esteso quasi come la fronte, la superficie dorsale è liscia o finemente granulata, le regioni non distinte; nel Terziario del Veneto la famiglia è rappresentata da: Archaeotetra lessinea, Braggitrapezia alessandroi De Angeli & Caporiondo, 2020, Eomaldivia lessinea De Angeli & Ceccon, 2017, E. trispinosa, Paratetralia convexa, P. sulcata (De Angeli et al., 2019; De Angeli & Caporiondo, 2020). Risulta abbastanza evidente che basandosi esclusivamente sulle caratteristiche del carapace non è facile attribuire un resto fossile ad una piuttosto che ad un’altra di queste ultime due famiglie come avevano correttamente rilevato Schweitzer (2005) e Karasawa & Schweitzer (2006) evidenziando come le specie fossili abbiano spesso caratteristiche intermedie e quindi ritenendo di attribuirle tutte ai Trapeziidae (vedi anche Beschin et al., 2007); in lavori successivi tuttavia alcune specie sono state inserite tra i Tetraliidae (Schweitzer et al., 2010, De Angeli & Ceccon , 2013b, Beschin et al. 2016, 2018, 2019). Sarebbe opportuno un più generale lavoro di approfondimento sull’argomento, anche se Nget al. (2008) sostengono che la mancanza dei caratteri diagnostici legati all’addome e alle parti ventrali compromette seriamente una corretta classificazione.
I carapaci attribuiti a Vicetitrapezia exaltissimo gen. nov., sp. nov. pur avendo una conformazione complessiva che li avvicina ai Trapeziidae secondo i caratte
ri indicati da Schweitzer (2005, tab. 2) come la fronte molto ampia che occupa quasi del tutto i margini anteriori, orbite rivolte antero-lateralmente, superficie dorsale non suddivisa in regioni e nel complesso liscia, presentano però delle caratteristiche che non consentono di attribuirli a tale famiglia e precisamente una curvatura in senso longitudinale molto accentuata e margini laterali con una evidente concavità nel tratto mediano per cui viene proposta l’istituzione della nuova famiglia Vicetitrapeziidae fam. nov.
Genere Vicetitrapezia gen. nov.
Specie tipo: Vicetitrapezia exaltissimo sp. nov.
Origine del nome: Vicetitrapezia (f.); nome composto da Vicetia, antica denominazione della città di Vicenza nel cui territorio è stata rinvenuta la serie tipo, e Trapezia Latreille, 1828, genere da cui deriva la denominazione della superfamiglia in cui il nuovo genere è inserito.
Diagnosi. La stessa della specie tipo.
Diagnosis. As the type species.
Vicetitrapezia exaltissimo sp. nov.
(figg. 64-66)
Serie tipica. Olotipo: esemplare MCZ 6194 (fig. 65). Paratipi: esemplare MCZ 6195, MCZ 6196, MCZ 6197, MCZ 6198, MCZ 6199, MCZ 6200 (fig. 66), MCZ 6201, MCZ 6202.
Località tipo: Altissimo, Vicenza
Livello tipo: Ypresiano (Eocene inferiore).
Origine del nome: exaltissimo (lat.) = da Altissimo; con riferimento alla località di rinvenimento della serie tipo.
Materiale e dimensioni. Nove carapaci: (VdP) MCZ 6197 L: 3,4 l: 3,0; MCZ 6196 L: 4,5 l: 3,8; MCZ 6195 L: 5,6 l: 5,1; MCZ 6194 L: 8,5 l: 7,4; MCZ 6200 l: 3,6; MCZ 6199 l: 5,9; MCZ 6198 l: 6,3; MCZ 6201, MCZ 6202. MCZ 6202 associato a Scutata eocenica MCZ 6050.
Diagnosi. Carapace subottagonale, poco più largo che lungo, molto convesso in senso longitudinale. Fronte molto ampia, rivolta verso il basso, con margine bilobo, debolmente ondulato. Orbite abbastanza grandi, poco profonde rivolte antero-lateralmente; dente extraorbitale robusto e appuntito. Margini antero-laterali nel primo tratto diritti e paralleli; poi con concavità semicircolare; margini postero-laterali diritti e nettamente convergenti; margine posteriore diritto e più stretto della fronte. Superficie dorsale non suddivisa in regioni, nel complesso liscia finemente punteggiata.
Diagnosis. Suboctagonal carapace, weakly wider than long, longitudinally strongly convex. Front very broad, downward folded, with bilobed, weakly wavy margin. Orbits quite large, shallow, anterolateral directed; outer orbital tooth stout and pointed. Anterolateral margins straight and parallel in the first part, then with semicircular concavity; posterolateral margins straight and strongly convergent; posterior margin straight and narrower than front. Dorsal surface not divided into regions, smooth with tiny dots.
Descrizione. Carapace subottagonale, poco più largo che lungo. Risulta molto convesso in senso longitudinale: l’area fronto-orbitale è molto ripida, quella
comprendente le regioni gastrica, cardiaca e branchiali anteriori è regolarmente convessa, mentre le regioni posteriori del carapace, con quella intestinale molto sviluppata, sono fortemente inclinate. La fronte è estremamente ampia e rivolta verso il basso; presenta un margine con andamento debolmente ondulato: è suddivisa infatti in due leggeri, grandi lobi separati da un debole ampio seno mediano. Ai lati si sviluppano le orbite rivolte antero-lateralmente, che sono abbastanza grandi ma non molto profonde; l’angolo intraorbitale non è distinto dalla fronte, il margine sovraorbitale è continuo e finemente bordato; il dente extraorbitale è invece robusto e appuntito. Le aree epatiche sono depresse. I margini antero-laterali sono nel primo tratto diritti e subparalleli; disegnano poi una profonda concavità semicircolare; quelli postero-laterali sono diritti e nettamente convergenti fino al margine posteriore che è abbastanza lungo e rettilineo pur essendo decisamente più stretto della fronte. Il dorso non è suddiviso in regioni; solo alcune sono riconoscibili come deboli rigonfiamenti e si intuiscono i solchi cardiobranchiali. La superficie è nel complesso liscia ma finemente punteggiata.
Osservazioni. I carapaci oggetto di studio sono inseriti tra i Trapezioidea per la fronte estremamente larga, le orbite poste quasi a livello dell’angolo laterale, la superficie dorsale non suddivisa in regioni. Alcuni caratteri però non si riscontrano in nessuna delle specie viventi attribuite alle tre famiglie Domeciidae, Tetraliidae e Trapeziidae secondo quanto proposto da Castro et al. (2004) né tra quelle fossili elencate da Schweitzer et al. (2010), De Angeli et al. (2019) e De Angeli & Caporiondo (2020). Si tratta della particolare curvatura longitudinale del dorso e la struttura dei margini laterali che presentano una concavità semicircolare che rendono per il momento unica questa specie. Un certo grado di convessità longitudinale nel carapace si riscontra anche in Paratetralia convexa ma localizzata essenzialmente nella parte anteriore. Alcune somiglianze si trovano con Archaeotetra lessinea nella conformazione della fronte e con Scutata eocenica per il grande sviluppo della parte posteriore del carapace.
Superfamiglia Xanthoidea MacLeay, 1838
Famiglia Panopeidae Ortmann, 1893
Genere Bittnereus Beschin, Busulini, De Angeli & Tessier, 2007
Specie tipo: Panopaeus vicentinus (sic!) Bittner, 1875
Bittnereus vicentinus (Bittner, 1875)
(fig. 67)
1875 Panopaeus Vicentinus Bittner, p. 33, t. 2, f. 7.
2006 Panopaeus vicentinus – De Angeli & Garassino, p. 69 (cum syn.).
2007 Bittnereus vicentinus – Beschin et al., p. 44, t. 6, ff. 2a, b, 5a, b, 6.
2016 Bittnereus vicentinus – Beschin et al., p. 115, t. 14, f. 6.
Materiale e dimensioni. Un carapace: (VC) MCZ 6008 L: 5,8.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca – Verona) (Beschin et al., 2007, 2016). Eocene medio: Veneto (San Giovanni Ilarione – Verona) (Bittner, 1875). Eocene del Friuli (Buttrio – Udine) (Dainelli, 1915).
Genere Laevicarcinus Lőrenthey in Lőrenthey & Beurlen, 1929
Specie tipo: Laevicarcinus egerensis Lőrenthey in Lőrenthey & Beurlen, 1929
Laevicarcinus serratus Beschin, Busulini & Tessier, 2016
(figg. 68, 69)
2016 Laevicarcinus serratus Beschin, Busulini & Tessier, p. 119, fig. 58, t. 15, f. 4.
2019 Laevicarcinus serratus – Beschin et al., p. 109, fig. 90.
Materiale dimensioni. Un carapace: (VC) MCZ 6011 l > 5,7; (VP) MCZ 6012 L > 6,3.
Osservazioni. Anche se gli esemplari considerati sono in cattivo stato di conservazione la loro attribuzione è sicura. MCZ 6011 è fortemente decorticato ma la forma generale del carapace, le spine laterali e la suddivisione in regioni del dorso che appaiono rigonfie sono confrontabili con quelle di L. serratus (poco evidente solo il lobo metagastrico); MCZ 6012 conserva solo le regioni anteriori ma conformi alla specie sono la fronte, i denti antero-laterali finemente denticolati e tutte le regioni osservabili.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Laevicarcinus sp.
(fig. 70)
Materiale e dimensioni. Un carapace incompleto lungo i margini laterali: (VP) MCZ 6009 L > 7,2.
Descrizione. Il carapace è poco convesso, incompleto ai lati, con superficie quasi del tutto decorticata. La fronte è quasi diritta, costituita da due lamelle. Il margine orbitale è concavo, nettamente separato da quello frontale. I margini laterali e quello posteriore non sono conservati. Le regioni sono leggermente rilevate e ben definite da solchi. Si osservano i lobi epigastrici e quelli protogastrici che risultano ampi, subovali, con debole carena anteriore. Il corpo del lobo mesogastrico è pentagonale, con un processo anteriore stretto e lungo che non raggiunge la fronte. La regione cardiaca è poco più larga del lobo mesogastrico, delimitata da solchi branchio-cardiaci ben incisi. Regioni epatiche triangolari con debole carena trasversale; regioni branchiali suddivise in lobi ma non complete; si intuisce una leggera carena sul lobo epibranchiale.
Osservazioni. Il carapace nella parte anteriore è molto simile a Laevicarcinus lioyi Beschin, Busulini, De Angeli & Tessier, 2007, specie diffusa in accumuli organogeni eocenici del Veneto (Beschin et al., 2007, 2015, 2016, 2019; Tessier et al., 2011). La regione cardiaca appare però più stretta, mentre le caratteristiche delle regioni branchiali non sono del tutto leggibili. Per questi motivi si è preferito mantenere aperta la determinazione specifica.
Genere Panopeus H. Milne Edwards, 1834
Specie tipo: Panopeus herbsti H. Milne Edwards, 1834
Panopeus incisus Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 71)
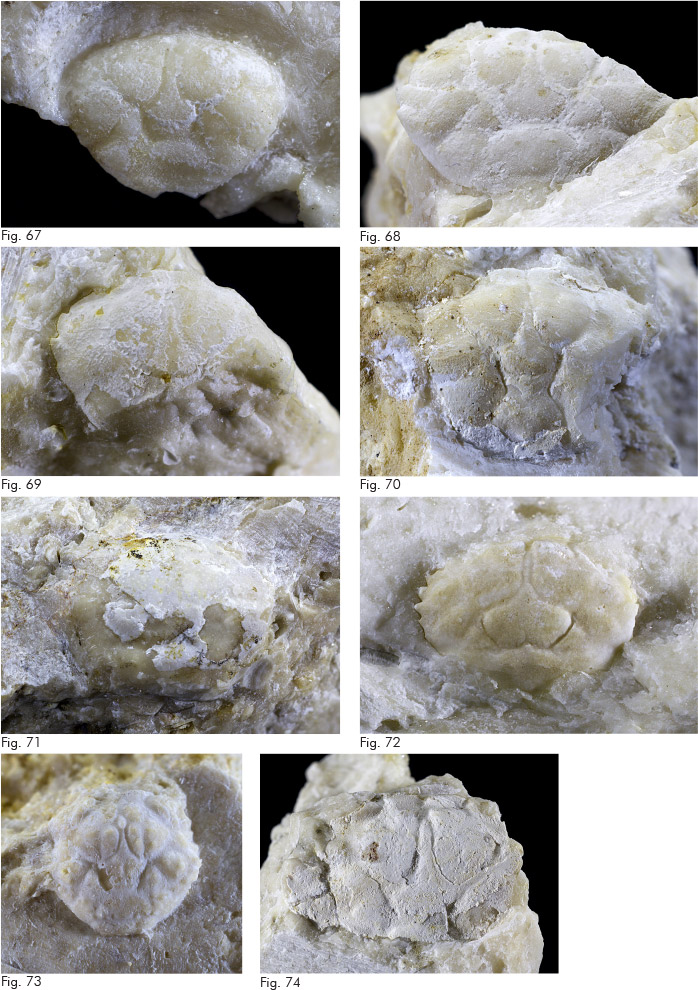
Fig. 68. Laevicarcinus serratus Beschin, Busulini & Tessier, 2016, es. MCZ 6011 (misura parziale l: 5,7), carapace, visione dorsale.
Fig. 69. Laevicarcinus serratus Beschin, Busulini & Tessier, 2016, es. MCZ 6012 (misura parziale L: 6,3), carapace, visione dorsale.
Fig. 70. Laevicarcinus sp., es. MCZ 6009 (misura parziale L: 7,2), carapace, visione dorsale.
Fig. 71. Panopeus incisus Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6013 (misura parziale l: 5,3), carapace, visione dorsale.
Fig. 72. Panopeus postalensis Beschin Busulini & Tessier, 2016, es. MCZ 6014 (L: 5,3), carapace: visione dorsale.
Fig. 73. Etisus arduinoi Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 5992 (L: 3,0 l: 2,9), carapace: visione dorsale.
Fig. 74. Haydnella maladensis Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6016 (misura parziale L: 15,2), carapace: visione dorsale.
2007 Panopeus incisus Beschin, Busulini, De Angeli & Tessier, p. 47, t. 6, ff. 3, 4.
2016 Panopeus incisus – Beschin et al., p. 120, t. 5, f. 5.
2018 Panopeus incisus – Beschin et al., p. 190, fig. 114a,b.
Materiale e dimensioni. Un carapace: (CZ) MCZ 6013 l > 5,3.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo -Vicenza; Bolca – Verona) (Beschin et al., 2007, 2016); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza) (Beschin et al., 2018).
Panopeus postalensis Beschin Busulini & Tessier, 2016
(fig. 72)
2016 Panopeus postalensis – Beschin Busulini & Tessier, p. 121, fig. 59, t. 15, ff. 6-8.
2019 Panopeus postalensis – Beschin et al., p. 111, fig. 93.
Materiale e dimensioni. Un carapace: (VdP) MCZ 6014 L: 5,3.
Distribuzione. Eocene inferiore: Veneto (Monte Postale, Altissimo – Vicenza) (Beschin et al., 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Famiglia Xanthidae MacLeay, 1838
Genere Etisus H. Milne Edwards, 1834
Specie tipo: Cancer dentatus Herbst, 1785
Etisus arduinoi Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 73)
2007 Etisus arduinoi Beschin, Busulini, De Angeli & Tessier, p. 58, t. 8, fig. 9-11.
2016 Etisus arduinoi – Beschin et al., p. 128, t. 16, f. 7 (cum.syn.).
2018 Etisus arduinoi – Beschin et al., p. 194, fig. 121.
Materiale e dimensioni. Un carapace: (CC) MCZ 5992 L: 3,0 l: 2,9.
Osservazioni. La suddivisione in regioni del carapace permette di avvicinarlo a Etisus arduinoi, specie fossile di ambiente corallino diffusa nel Veneto occidentale; analoghe risultano infatti la fronte larga, depressa, costituita da due lamelle, le regioni ben definite, rigonfie e tubercolate con lobi protogastrici caratterizzati da due rilievi longitudinali tubercolati e lobo mesogastrico pentagonale con processo anteriore stretto e allungato con tubercolo apicale, quelle branchiali suddivise da un solco subtrasversale e da due solchi obliqui. Si osserva che il carapace presenta lunghezza e larghezza quasi equidimensionali, mentre in tutti gli esemplari finora attribuiti alla specie la larghezza supera nettamente la lunghezza. Va notato, comunque, che il carapace oggetto di studio è decisamente più piccolo rispetto a tutti gli altri esemplari noti e che in generale quelli più grandi hanno un rapporto L/l maggiore di quelli più piccoli. Questa tendenza si osserva in altre specie fossili come ricordato da Beschin et al. (2017). Il resto rinvenuto ad Altissimo potrebbe dunque essere un esemplare con caratteristiche giovanili.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca e Vestenanova – Verona); Eocene superiore (Campolongo di Val Liona – Vicenza) (Beschin et al., 2007, 2015, 2016, 2018; Tessier et al., 2011).
Genere Haydnella Müller, 1984
Specie tipo: Haydnella steiningeri Müller, 1984
Haydnella maladensis Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 74)
2007 Haydnella maladensis Beschin, Busulini, De Angeli & Tessier, p. 59, t. 9, f. 1.
2019 Haydnella maladensis – Beschin et al., p. 113, fig. 96 (cum syn.).
Materiale e dimensioni. Il carapace incompleto: (VdP) MCZ 6016 L > 15,2.
Osservazioni. Nonostante sia estremamente incompleto, l’esemplare può essere riferito alla specie per i lobi epigastrici triangolari molto evidenti, i robusti denti antero-laterali, il solco ondulato a forma di giogo che separa le regioni anteriori da quelle posteriori del carapace, la presenza di abbondante granulazione sul dorso.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Braggi, Bolca – Verona) (Beschin et al., 2007, 2015, 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Genere Phlyctenodes A. Milne Edwards, 1862
Specie tipo: Phlyctenodes tuberculosus A. Milne Edwards, 1862
Phlyctenodes tuberculosus A. Milne Edwards, 1862
(fig. 75)
1862 Phlyctenodes tuberculosus A. Milne Edwards, p. 193, t. 7, f. 2.
2009a Phlyctenodes tuberculosus – Beschin et al., p. 19, t. 3, f. 6.
2018 Phlyctenodes tuberculosus – Beschin et al., p. 194, fig. 123 (cum syn.).
2019 Phlyctenodes tuberculosus – Beschin et al., p. 113, fig. 98.
Materiale e dimensioni. Un carapace: (VC) MCZ 6027 l: 4,9.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016); Eocene medio: Francia sudoccidentale (“terrain nummulitique” di Hastingues – Landes) (Milne Edwards, 1862); Eocene superiore: Veneto (San Feliciano e Campolongo di Val Liona – Vicenza; Parona – Verona) (Busulini et al., 2006; Beschin et al., 2009a, 2018, 2019).
Phlyctenodes edwardsi Beschin, Busulini & Tessier, 2016
(fig. 76)
2016 Phlyctenodes edwardsi Beschin, Busulini & Tessier in Beschin et al., p.133, fig. 65, t. 17, ff. 4 a, b.
Materiale e dimensioni. Due carapaci: (VP) MCZ 6029 l: 2,5; MCZ 6028 l > 5,8.
Distribuzione. Eocene inferiore: Veneto (Rama di Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Phlyctenodes multituberculatus Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 77)
2000 Phlyctenodes cf. steinmanni – Beschin et al., p. 8, t. 2, f. 4
2007 Phlyctenodes multituberculatus Beschin, Busulini, De Angeli & Tessier, p. 62, t. 10, ff. 1-4.
2011 Phlyctenodes multituberculatus – Tessier et al., p. 219, f. 3.11.
2015 Phlyctenodes multituberculatus – Beschin et al., p. 93, t. 7, f. 3.
2016 Phlyctenodes multituberculatus – Beschin et al., p. 134, t. 17, f. 6.
2019 Phlyctenodes multituberculatus – Beschin et al., p. 114, fig. 101.
Materiale e dimensioni. Nove carapaci: (CC) MCZ 6019 L: 6,5 l: 5,1; MCZ 6018 L >7,9; MCZ 6020 l: 6,4; (VC) MCZ 6021 L: 7,7; MCZ 6026; (VdP) MCZ 6022; (VP) MCZ 6024 L: 6,9 l: 4,6; MCZ 6023 L: 8,2; MCZ 6025.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino – Vicenza; Bolca, Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Genere Speocarcinus Stimpson, 1859
Specie tipo: Speocarcinus carolinensis Stimpson, 1859
Speocarcinus latus Beschin, Busulini & Tessier, 2016
(fig. 78)
2016 Speocarcinus latus Beschin, Busulini & Tessier in Beschin et al., p. 135, fig. 66, t. 17, ff. 7 a, b.
2019 Speocarcinus latus – Beschin et al., p. 114, fig. 104.
Materiale e dimensioni. Un carapace decorticato: (VdP) MCZ 6048 L: 9,0 l: 6,2.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Xanthoidea incertae sedis sensu Schweitzer et al. (2010)
Genere Muelleroplax Schweitzer, Feldmann, Karasawa, Garassino & Schweigert, 2010
Specie tipo: Eoplax minima Müller & Collins, 1991.
Muelleroplax cfr. minuscula (Beschin, Busulini, De Angeli & Tessier, 2007)
(figg. 79, 80)
2007 Eoplax minuscula Beschin, Busulini, De Angeli & Tessier, p. 66, t. 10, f. 7.
Materiale e dimensioni. Due carapaci: (VdP) MCZ 6204 L: 6,2 l > 5,0; (VP) MCZ 6205 l: 5,3.
Osservazioni. Gli esemplari corrispondono bene a Muelleroplax minuscula, specie presente in biocostruzioni dell’Eocene inferiore nel Vicentino (contrada Gecchelina di Monte di Malo) e nel Veronese (Bolca e Vestenanova) (Beschin et al., 2007, 2015, 2016). L’incertezza nella attribuzione dipende dalla presenza di due fessure lungo il margine orbitale e dalla fronte un po’ più avanzata; si rileva comunque la non perfetta conservazione di queste parti dei carapaci rinvenuti ad Altissimo.
Genere Pilumnomimus Müller & Collins, 1991
Specie tipo: Pilumnomimus planidentatus Müller & Collins, 1991
Pilumnomimus planidentatus Müller & Collins, 1991
(fig. 81)
1991 Pilumnomimus planidentatus Müller & Collins, p. 75, t. 5, ff. 5, 8, 11, fig. 4h.
2007 Pilumnomimus cf. planidentatus – Beschin et al., p. 52. t. 7, f. 9.
2016 Pilumnomimus planidentatus – Beschin et al., p. 138, t. 18, f. 2.
2019 Pilumnomimus planidentatus – Beschin et al., p. 115, fig. 106.
Materiale e dimensioni. Due carapaci: (CZ) MCZ 6030 L: 7,8 l: 5,3; (VC) MCZ 6031 L: 3,4 l: 2,5. MCZ 6030 associato a Corallioplax exigua MCZ 6039.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca – Verona ) (Beschin et al., 2007, 2016); Eocene superiore: Ungheria (Müller & Collins, 1991), Veneto (Parona – Verona) (Beschin et al., 2019).
Genere Prochlorodius Müller & Collins, 1991
Specie tipo: Prochlorodius ellipticus Müller & Collins, 1991
Prochlorodius ellipticus Müller & Collins, 1991
(fig. 82)

Fig. 76. Phlyctenodes edwardsi Beschin, Busulni & Tessier, 2016, es. MCZ 6028 (misura parziale l: 5,8), carapace: visione dorsale.
Fig. 77. Phlyctenodes multituberculatus Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6019 (L: 6,5 l: 5,1), carapace: visione dorsale.
Fig. 78. Speocarcinus latus Beschin, Busulini & Tessier, 2016, es. MCZ 6048 (L: 9,0 l: 6,2), carapace: visione dorsale.
Fig. 79. Muelleroplax cfr. minuscula (Beschin, Busulini, De Angeli & Tessier, 2007), es. MCZ 6204 (L: 6,2), carapace: visione dorsale.
Fig. 80. Muelleroplax cfr. minuscula (Beschin, Busulini, De Angeli & Tessier, 2007), es. MCZ 6205 (l: 5,3), carapace: visione dorsale.
Fig. 81. Pilumnomimus planidentatus Müller & Collins, 1991, es. MCZ 6030 (L: 7,8 l: 5,3), carapace: visione dorsale.
Fig. 82. Prochlorodius ellipticus Müller & Collins, 1991, es. MCZ 6045 (l: 6,0), carapace:visione dorsale.
1898 Neptocarcinus millenaris – Lőrenthey, p. 69, t. 4, f. 4 (non f. 3).
1929 Neptocarcinus millenaris – Lőrenthey & Beurlen, p. 216, t. 10, f. 4 (non f. 3).
1991 Prochlorodius ellipticus Müller & Collins, p. 78, figg. 4i, j, t. 5, f. 11, t. 6, ff. 4, 7, 17.
2016 Prochlorodius ellipticus – Beschin et al., p. 140, t. 18, f. 5 (cum syn.).
2018 Prochlorodius ellipticus – Beschin et al., p. 196, figg. 131a, b.
2019 Prochlorodius ellipticus – Beschin et al., p. 115, figg. 107, 108.
Materiale e dimensioni. Sei carapaci: (CC) MCZ 6033 l: 4,7; MCZ 6034; (CZ) MCZ 6037 l: 8,5; MCZ 6036 l: 8,9; (VC) MCZ 6042 L: 5,7; (VdP) MCZ 6045 l: 6,0.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, San Pietro Mussolino, Altissimo – Vicenza; Bolca, Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016; Tessier et al., 2011); Eocene superiore: Ungheria (Lőrenthey, 1898; Lőrenthey & Beurlen, 1929; Müller & Collins, 1991) e Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Superfamiglia Eriphioidea MacLeay, 1838
Famiglia Pseudoziidae Alcock, 1898
Genere Ramozius Beschin, Busulini & Tessier, 2016
Specie tipo: Ramozius punctatus Beschin, Busulini & Tessier, 2016
Ramozius punctatus Beschin, Busulini & Tessier, 2016
(fig. 83)
2016 Ramozius punctatus Beschin, Busulini & Tessier in Beschin et al., p. 141, fig. 69, t. 18, ff. 6 a, b.
Materiale e dimensioni. Un carapace: (VP) MCZ 6203 L: 16,0.
Osservazioni. Il carapace, incompleto anteriormente, è decorticato e permette di osservare strutture che non erano evidenti nell’olotipo (Beschin et al.,
2016) che invece conservava la cuticola intatta e in particolare: le inserzioni dei muscoli epimerali, una leggera cresta epibranchiale a convessità anteriore, tre tubercoli cardiaci, alcuni tubercoli ai lati del lobo mesogastrico.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Superfamiglia Goneplacoidea MacLeay, 1838
Famiglia Euryplacidae Stimpson, 1871
Genere Alponella Beschin, Busulini & Tessier, 2016
Specie tipo: Alponella paleogenica Beschin, Busulini & Tessier, 2016.
Alponella paleogenica Beschin, Busulini & Tessier, 2016
(fig. 84)
2016 Alponella paleogenica Beschin, Busulini & Tessier in Beschin et al., p. 143, fig. 70, t. 19, ff. 1 a, b.
2019 Alponella paleogenica – Beschin et al., p. 117, fig. 110.
Materiale e dimensioni. Un carapace: (VC) MCZ 6214 l: 2,2.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Genere Corallioplax Beschin, Busulini & Tessier, 2016
Specie tipo: Corallioplax exigua Beschin, Busulini & Tessier, 2016
Corallioplax exigua Beschin, Busulini & Tessier, 2016
(fig. 85)
2015 Branchioplax sp. 1 – Beschin et al., p. 100, t. 8, f. 4.
2016 Corallioplax exigua Beschin, Busulini & Tessier in Beschin et al., p. 144, fig. 71, t. 19, ff. 2 a, b, 3.
2019 Corallioplax exigua – Beschin et al., p. 117, fig. 112.
Materiale e dimensioni. 10 carapaci: (CC) MCZ 6032 l: 6,9; (CZ) MCZ 6035 L: 5,5 l: 4,1; MCZ 6038 l: 6,3; MCZ 6039; (VC) MCZ 6041 L: 6,7; MCZ 6040 L: 8,5; MCZ 6043; MCZ 6044; (VdP) MCZ 6046 l: 4,6; (VP) MCZ 6047 l: 4,8.
MCZ 6039 associato a Pilumnomimus planidentatus MCZ 6030; MCZ 6044 associato a Paratetralia convexa MCZ 6092.
Distribuzione. Eocene inferiore: Veneto (Vestenanova, Altissimo – Vicenza; Bolca – Verona) (Beschin et al., 2015, 2016); Eocene superiore: Veneto (Parona – Verona) (Beschin et al., 2019).
Famiglia Mathildellidae Karasawa & Kato, 2003
Genere Branchioplax Rathbun, 1916
Specie tipo: Branchioplax washingtoniana Rathbun, 1916 (= Neopilumnoplax hannibalanus Rathbun, 1926)
Branchioplax cordata Beschin, Busulini & Tessier, 2016
(fig. 86)
2016 Branchioplax cordata Beschin, Busulini & Tessier in Beschin et al., p. 147, fig. 73, t. 19, ff. 5, 6.
Materiali e dimensioni. Tre carapaci: (CZ) MCZ 6208 L: 7,6 l: 6,1; (VC) MCZ 6209 L: 5,0 l: 3,9; (VP) MCZ 6215 l: 5,3. MCZ 6209 associato a Archaeotetra lessinea MCZ 6064.
Distribuzione. Eocene inferiore: Veneto (Bolca – Verona; Altissimo – Vicenza) (Beschin et al., 2016).
Branchioplax parva Beschin, Busulini, De Angeli & Tessier, 2007
(fig. 87)
2007 Branchioplax parva Beschin, Busulini, De Angeli & Tessier, p. 36, t. 5, ff. 3, 4.
2016 Branchioplax parva – Beschin et al., p. 148, t. 20, f. 1 (cum syn.).
2018 Branchioplax parva – Beschin et al., p. 201, figg. 139, 140.
2019 Branchioplax parva – Beschin et al., p. 119, fig. 115.
Materiale e dimensioni. Un carapace: (VC) MCZ 6213 L: 4,1; è associato a Dromiopsis marginospinosa MCZ 5938.
Distribuzione. Eocene inferiore: Veneto (contrada Gecchelina di Monte di Malo, Altissimo – Vicenza; Bolca e Vestenanova – Verona) (Beschin et al., 2007, 2015, 2016); Eocene superiore: Veneto (Campolongo di Val Liona – Vicenza; Parona – Verona) (Beschin et al., 2018, 2019).
Genere Clampethildella gen. nov.
Specie tipo: Clampethildella spinosa sp. nov.
Origine del nome: Clampethildella (f.) è ottenuto dalla fusione di Clampus, antica denominazione del torrente Chiampo, e Mathildella Guinot & Richer de Forges, 1981, genere tipo della famiglia in cui il nuovo genere è inserito.
Diagnosi. La stessa della specie tipo.
Diagnosis. As the type species.
Clampethildella spinosa sp. nov.
(fig. 88, 89)
Serie tipica. Olotipo: esemplare MCZ 6206 (fig. 89). Paratipo: esemplare MCZ 6207.
Località tipo: Altissimo, Vicenza
Livello tipo: Ypresiano (Eocene inferiore).
Origine del nome: spinosa (lat.) = ricca di spine, con riferimento alle numerose piccole spine che ornano i margini del carapace.
Materiale e dimensioni. Due carapaci: (CZ) MCZ 6206 l: 4,6; (VP) MCZ 6207 l: 3,6.
Diagnosi. Carapace subesagonale, più largo che lungo, moderatamente convesso. Fronte ampia, diritta con incisione mediana; orbite grandi con una fessura; margini antero-laterali corti, convessi con due denti (escluso il dente extraorbi
tale); margini postero-laterali più lunghi, convessi e convergenti; margine posteriore ampio. Margini anteriori e parte di quelli postero-laterali con minute spine. Regioni poco definite da deboli solchi; superficie epatica ed epibranchiale granulata. Regione cardiaca trilobata, rilevata.
Diagnosis. Carapace subhexagonal, wider than long, mildly convex. Front broad, straight with median notch; orbits large with one fissure; anterolateral margins short, convex with two teeth (excluding outer orbital tooth); posterolateral margins longer, convex and convergent; posterior margin broad. Anterior margins and part of posterolateral ones with tiny spines. Regions poorly defined by weak furrows; hepatic and epibranchial surface granulate. Cardiac region trilobate, inflated.
Descrizione. Carapace subesagonale, più largo che lungo, relativamente convesso in entrambe le sezioni e con la massima larghezza nel terzo anteriore in corrispondenza dell’ultimo dente antero-laterale. Margine fronto-orbitale abbastanza esteso. Fronte poco sporgente oltre le orbite, con margine quasi diritto ma con leggera incisione mediana. Orbite relativamente grandi, delimitate dalla fronte da un piccolo dente preorbitale mentre quello extraorbitale è poco evidente; margine sopraorbitale concavo, rigonfio, interessato da una corta fessura. Margini antero-laterali corti e convessi, con due denti poco sporgenti ed appuntiti che si sviluppano presso l’angolo laterale; dal seno che li separa si sviluppa un solco che interessa per un certo tratto il dorso del carapace delimitando posteriormene le regioni epatiche. I margini postero-laterali sono più sviluppati dei precedenti, convessi e convergenti; margine posteriore largo, quasi diritto segnato da un bordino rilevato, con rientranze coxali poco profonde. Tutti i margini anteriori e la parte iniziale di quelli postero-laterali sono ornati da minute spine. Le regioni del dorso sono in parte delimitate da deboli solchi. La regione frontale è nel complesso piatta mentre i lobi epigastrici sono un po’ rilevati anteriormente. I lobi protogastrici sono relativamete stretti, delimitati solo nella parte posteriore dai rami laterali del solco cervicale. Il lobo meso-metagastrico è ribassato ma chiaramente delimitato; presenta corpo ovale e processo anteriore piuttosto robusto con apice triangolare oltre i lobi epigastrici. La regione cardiaca è trilobata, evidente e decisamente rilevata; la regione intestinale è piccola. Le regioni epatiche sono abbastanza estese, parzialmente delimitate posteriormente; portano una leggera cresta obliqua. Le regioni branchiali sono ampie, segnate da un modesto solco branchiale trasversale con leggera concavità posteriore. La superficie dorsale mostra granuli maggiormente evidenti sulle aree epatiche posteriori ed epibranchiali.
Osservazioni. Gli esemplari analizzati ricordano i rappresentanti del genere Branchioplax per la forma generale del carapace che si presenta subesagonale e moderatamente convesso, per la conformazione della fronte, per i margini antero-laterali relativamente brevi e dentati, per i margini postero-laterali convessi e per la suddivisione in regioni definite da solchi poco incisi. Le specie attribuite a tale genere sono diffuse dall’Eocene inferiore all’Oligocene in varie parti del mondo (Europa, Senegal, Giappone, Stati Uniti) (Beschin et al., 2007, 2016) e sono presenti anche in Veneto con le specie eoceniche B. albertii De Angeli & Beschin, 2002, B. cordata, B. parva, B. sulcata Müller & Collins, 1991, Branchioplax sp. 2 di Beschin et al. (2015) e la oligocenica B. rossii De Angeli & Beschin, 2008. Gli esemplari rinvenuti ad Altissimo si distinguono però da tutte per la presenza di un’unica fessura sul margine sovraorbitale (anziché 2), due denti antero-laterali (anziché 3) e la presenza di minute spine sui margini anteriori e la parte iniziale di quelli postero-laterali. Queste differenze giustificano l’istituzio
ne di un nuovo genere e di una nuova specie all’interno dei Mathildellidae. Tra i taxa inclusi in questa famiglia, margini anteriori del carapace ornati da minute spine, due denti sui margini antero-laterali e margini orbitali con una fessura si ritrovano in Platypilumnus gracilipes Alcock, 1894, che vive nei mari delle Filippine; la forma generale del carapace che è proporzionalmente più allungato con fronte più sporgente e regioni posteriori più marcate allontanano tuttavia questa specie da quella di nuova istituzione (Nget al., 2008).
Chelipedi indeterminati
Vengono di seguito descritti resti di alcuni chelipedi isolati rinvenuti nei vari accumuli organogeni del territorio di Altissimo la cui attribuzione rimane incerta. Non essendo definiti dal punto di vista sistematico, questi esemplari non sono stati inseriti nella tabella 1 per non alterare i risultati degli indici statistici calcolati.
Chelipede 1 (fig. 90)
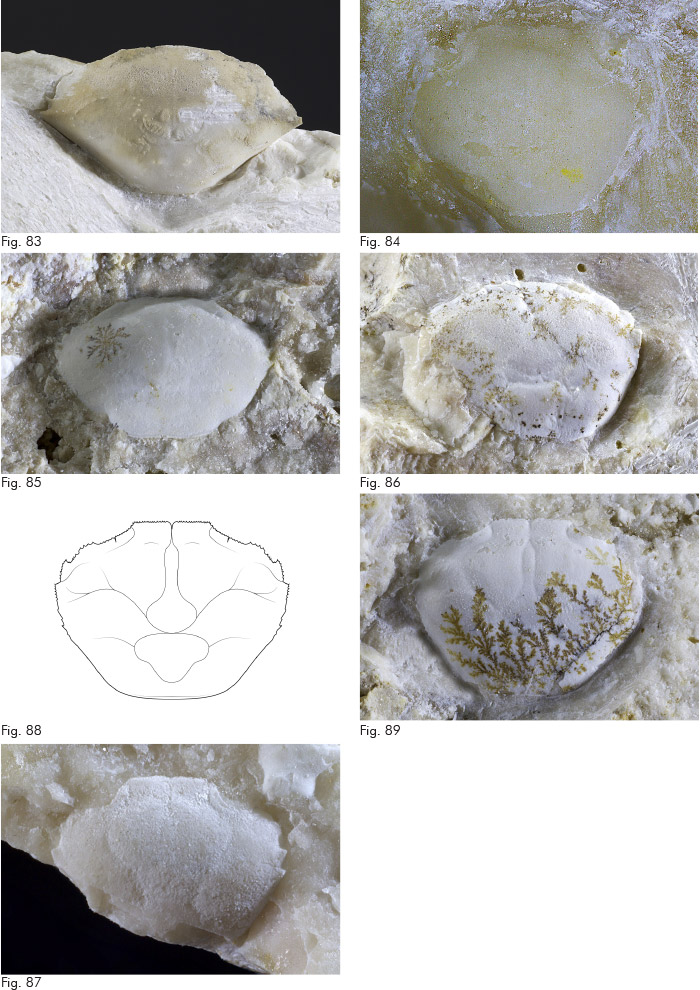
Fig. 84. Alponella paleogenica Beschin, Busulini & Tessier, 2016, es. MCZ 6214 (l: 2,2), carapace:visione dorsale.
Fig. 85. Corallioplax exigua Beschin, Busulini & Tessier, 2016, es. MCZ 6035 (L: 5,5 l: 4,1), carapace: visione dorsale.
Fig. 86. Branchioplax cordata Beschin, Busulini & Tessier, 2016, es. MCZ 6208 (L: 7,6 l: 6,1), carapace: visione dorsale.
Fig. 87. Branchioplax parva Beschin, Busulini, De Angeli & Tessier, 2007, es. MCZ 6213 (L: 4,1), carapace: visione dorsale.
Fig. 88. Clampethildella spinosa gen. nov., sp. nov., profilo schematico di carapace.
Fig. 89. Clampethildella spinosa gen. nov., sp. nov., olotipo, es. MCZ 6206 (l: 4,6), carapace: visione dorsale.
Materiale e dimensioni. Un propodo sinistro in visione esterna: (CC) MCZ 6217 lp: 7,7 h: 6,0.
Descrizione. Del propodo è conservato solamente il palmo che è subrettangolare un po’ allungato con margini superiore ed inferiore subparalleli ma leggermente convessi e sottolineati da un sottile bordo; quello inferiore è spinoso. La superficie è poco bombata, interessata da allineamenti trasversali subparalleli di piccole “squame” dirette distalmente. Articolazione carpo-propodiale subverticale e relativamente stretta.
Chelipede 2 (fig. 91)
Materiale e dimensioni. Un chelipede sinistro in visione esterna che conserva parte di mero, carpo, propodo e dattilo: (VdP) MCZ 6218 h: 8,5.
Descrizione. Propodo con palmo subrettangolare, poco allungato, con articolazione carpo-propodiale appena inclinata. Margine superiore con un grosso tubercolo prossimale collegato all’articolazione e una serie di denti; margine inferiore convesso e poi concavo alla base del dito fisso; margine distale con una profonda tacca superiore poi convesso e infine concavo; dito fisso corto, triangolare e appuntito, diretto un po’ in basso, con margine occlusale liscio; superficie fortemente abrasa. Dattilo robusto, falciforme con una convessità sulla parte prossimale del margine superiore. Carpo caratterizzato da numerosi denti.
Chelipede 3 (fig. 92)
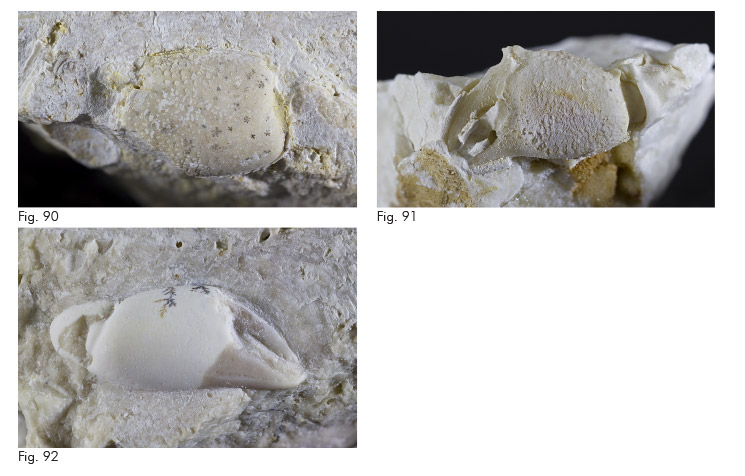
Fig. 91. Chelipede 2, es. MCZ 6218 (h propodo: 8,5), chelipede sinistro.
Fig. 92. Chelipede 3, es. MCZ 6219 (lp: 4,4 h: 4,1), propodo e dattilo destri.
Materiale e dimensioni. Tre chele in visione esterna: (CC) MCZ 6219 (destra) lp: 4,4 h: 4,1; MCZ 6220 (propodo sinistro) lp: 4,4 h: 4,1; (VC) MCZ 6221 (destra) h: 2,5.
Descrizione. Le due chele destre sono complete del dattilo e mostrano ancora parte del carpo in connessione anatomica. Propodo allungato con articolazione carpo-propodiale molto inclinata distalmente. Palmo ovale poco più lungo che alto con margini superiore ed inferiore convessi, lisci, nel complesso subparalleli; sugli angoli prossimali superiore ed inferiore si osservano dei rigonfiamenti collegati all’articolazione. Dopo un tratto concavo il margine inferiore continua nel dito fisso che è piuttosto lungo e appuntito con margine occlusale caratterizzato da alcuni denti. La superficie è liscia con una serie di pori setali allineati lungo il dito fisso. In MCZ 6220, con cuticola abrasa, si osservano delle porosità diffuse. Dattilo falciforme e liscio. Il carpo non è osservabile perché immerso nella matrice rocciosa.
Conclusioni
L’analisi della fauna di decapodi associati a coralli rinvenuta in vari ammassi organogeni affioranti nel comune di Altissimo (Vicenza) fornisce un nuovo utile contributo alla conoscenza di questo gruppo per l’Eocene inferiore.
I siti dove sono stati rinvenuti i crostacei si trovano a breve distanza dalle località oggetto di ricerca e studio negli anni precedenti. Queste delimitano un’area all’interno della quale si trova Altissimo da cui Bolca dista 4 km in direzione OSO, Vestenanova (Cava Braggi) 5,75 km SSO, San Pietro Mussolino (Cava Bertocchi) 2,80 km SSE, contrada Gecchelina di Monte di Malo 10,7 km NE mentre Monte Magrè di Schio 9,35 km NNE (Beschin et al., 2007, 2015, 2016, 2018, 2019; Tessier et al., 2011; De Angeli & Ceccon, 2012, 2013a,b, 2014, 2015, 2016a,b, 2017, 2020; De Angeli & Garassino, 2014; De Angeli & Caporiondo, 2016, 2020).
Si tratta di un ulteriore importante elemento a conferma dell’esistenza nell’Eocene inferiore di un’ampia laguna caratterizzata da acque basse, con numerose biocostruzioni ad alghe e coralli dove prosperavano i molluschi e una ricca e ben diversificata fauna di crostacei la cui composizione è ormai ben definita. Nei reperti oggetto di studio infatti sono state riconosciute quasi tutte le specie indicate da Beschin et al. (2017) come caratterizzanti l’associazione fossile delle altre località indagate e precisamente tra gli anomuri Lessinigalathea regalis e tra i brachiuri Dromiopsis paucigranosa, Kromtitis koberiformis, Cyamocarcinus angustifrons e C. budensis, Eotrachynotocarcinus airaghii, Galenopsis similis, Lobogalenopsis quadrilobata, Paratetralia convexa, Etisus arduinoi, Phlyctenodes multituberculatus e Prochlorodius ellipticus; non sono presenti solo Daira sicula (Di Salvo, 1933), Gecchelicarcinus lorigae Beschin, Busulini, De Angeli & Tessier, 2007 (è presente però G. zanderigoi) e Laevicarcinus lioyi (avente notevoli somiglianze con Laevicarcinus sp.).
Ma accanto a specie già note, i nuovi rinvenimenti hanno permesso di descrivere alcuni nuovi taxa: tre nuovi generi e specie, Cristinagalathea striata gen. nov., sp. nov. (Munidopsidae), Clampethildella spinosa gen. nov., sp. nov. (Mathildellidae) e Vicetitrapezia exaltissimo gen. nov., sp. nov.; per quest’ultimo si è ritenuto di creare una nuova famiglia di Trapezioidea, Vicetitrapeziidae fam. nov., tenuto conto delle caratteristiche uniche degli esemplari, che non permettevano un’utile inquadramento nelle famiglie note. È stato inoltre istituito Muellermunida gen. nov. (Munididae) per una migliore collocazione sistematica di Protomunida pentacantha e di Eumunida veronensis. Grazie al ritrovamento di un esemplare più completo rispetto all’olotipo viene fornita una diagnosi emendata di Paguristes vallionensis (Diogenidae). È presentato inoltre un esemplare di Etisus arduinoi con caratteristiche giovanili.
Dal punto di vista della abbondanza relativa (Ar) delle specie si osserva che quasi tutti i taxa sono presenti con un numero molto esiguo di esemplari e quindi con un valore di Ar molto basso, con l’eccezione di Paratetralia convexa (Ar = 0,310) seguita da un altro trapezide Archaeotetra lessinea (Ar = 0,059).
L’indice di biodiversità Shannon-Weaver (H’= 3,3) ha un valore confrontabile con quello già calcolato per le citate faune coeve di ambiente corallino recentemente descritte per il Vicentino e Veronese (Beschin et al., 2017).
Recenti studi hanno investigato la presenza dei crostacei negli ambienti a coralli, analizzando alcune zone della grande barriera corallina dell’Australia; l’analisi ha permesso di rilevare come le comunità a crostacei differiscano notevolmente a seconda del microhabitat: tra i coralli morti si registra la maggior abbondanza, con un valore 2 – 3 volte maggiore di quello relativo di quelli presenti all’interno delle colonie di coralli viventi (Kramer et al., 2014) per cui ci possono essere alcune differenze nelle associazioni faunistiche tra località anche molto vicine, come si riscontra negli affioramenti analizzati in questi anni.
È stato altresì sottolineato come, nonostante la grande diversità e abbondanza delle faune di invertebrati presenti negli ecosistemi corallini, l’importanza dei crostacei (che rappresentano circa il 20% di tutte le specie di invertebrati), sia stata spesso sottovalutata a causa della ridotta dimensione e della circostanza che vivono nascosti all’interno delle strutture coralline (Kramer et al., 2014).
Queste osservazioni confermano che i rinvenimenti di crostacei fossili associati a coralli sono fortemente influenzati dalle modalità delle ricerche proprio a causa della loro ridotta dimensione e delle loro abitudini di vita. I decapodi in particolare hanno un’importante funzione all’interno degli attuali ecosistemi delle barriere coralline; vivono in relazione simbiotica con i coralli ed anche con i pesci nei confronti dei quali è dimostrata una loro funzione di difesa dai predatori e di pulizia dai parassiti.
Tabella 1. Elenco dei crostacei fossili rinvenuti negli ammassi organogeni dell’Eocene inferiore affioranti nel comune di Altissimo (Vicenza). Sono riportati in colonne separate gli esemplari rinvenuti nelle varie località e precisamente: contrada Cocco (CC), contrada Zerbati (CZ), via Cero (VC), Valle dei Pini (VdP), via Pezzi (VP). Per ogni taxon è riportata la tipologia dei reperti (parte: c = carapace; p = propodo di chelipede; s = scudo) e l’indice di abbondanza relativa (Ar).
Ringraziamenti
Si ringraziano la Direzione e lo staff tecnico del Museo Civico “G. Zannato” di Montecchio Maggiore (Vicenza) per aver messo a disposizione per questo studio materiale conservato nelle collezioni dell’istituzione museale. Si ringraziano inoltre il prof. Ivano Rocchetti per l’esecuzione delle foto, il dott. Matteo Calvagno per l’esecuzione dei disegni illustrativi, il sig. Mariano Arsego per aver curato il disegno della localizzazione geografica dei siti indagati.
Un sentito grazie infine va al dott. Rodney M. Feldmann (Departement of Geology, Kent State University, USA) e al dott. Hiroaki Karasawa (Mizunami Fossil Museum, Mizunami, Giappone) per i preziosi consigli forniti e a due referee anonimi per la attenta lettura del testo e gli utili suggerimenti.
Bibliografia
AhyongT.S., Baba K., Macpherson E., Poore C.B. (2010) – A new classification of the Galatheoidea (Crustacea: Decapoda: Anomura). Zootaxa, 2676: 57-68.
Beschin C., Busulini A., De Angeli A., Tessier G., Ungaro S. (1991) – Due nuovi generi di Raninidae dell’Eocene del Veneto (Italia). Lavori Soc. ven. Sci. nat.,16:187- 212.
Beschin C., Busulini A., De Angeli A., Tessier G., Ungaro S. (2000) – The fauna of the Gecchelina quarry at Monte di Malo (Vicenza – Northern Italy): a preliminary study.“1st Workshop on Mesozoic and Tertiary decapod crustaceans”, Studi e ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato” – Montecchio Maggiore (Vicenza): 7-10.
Beschin C., Busulini A., De Angeli A., Tessier G. (2002) – Aggiornamento ai crostacei eocenici di cava “Main” di Arzignano (Vicenza – Italia settentrionale) (Crustacea, Decapoda). Studi e Ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato” – Montecchio Maggiore (Vicenza): 7-28.
Beschin C., De Angeli A., Checchi A., Zarantonello G. (2005) – Crostacei eocenici di Grola presso Spagnago (Vicenza – Italia settentrionale). Studi e Ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato” – Montecchio Maggiore (Vicenza), 12: 5-35.
Beschin C., De Angeli A., Checchi A., Mietto P. (2006) – Crostacei del Priaboniano di Priabona (Vicenza – Italia settentrionale). Lavori Soc. ven. Sci. nat., 31: 95-112.
Beschin C., Busulini A., De Angeli A., Tessier G. (2007) – I decapodi dell’Eocene inferiore di Contrada Gecchelina (Vicenza – Italia settentrionale) (Anomura e Brachyura). Mus. Archeol. Sci. nat. “G. Zannato”, Montecchio Maggiore (Vicenza): 76 pp.
Beschin C., Busulini A., Tessier G. (2009a) – The decapod crustaceans from the upper Eocene of Parona (Veronese Lessini – NE Italy). Studi e Ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato” – Montecchio Maggiore (Vicenza), 16: 5-22.
Beschin C., De Angeli A., Zorzin R. (2009b) – Crostacei fossili del Veneto: una inedita fauna eocenica dei Lessini orientali (Monte Serea di San Giovanni Ilarione, Verona). Boll. Mus. civ. St. nat. Verona, 33: 59-83.
Beschin C., De Angeli A., Checchi A., Zarantonello G. (2012) – Crostacei del giacimento eocenico di Grola presso Spagnago di Cornedo Vicentino (Vicenza, Italia settentrionale) (Decapoda, Stomatopoda, Isopoda). Mus. Archeol. Sci. Nat. “G. Zannato” – Montecchio Maggiore (Vicenza): 99 pp.
Beschin C., Busulini A., Tessier G. (2015) – Nuova segnalazione di crostacei associati a coralli nell’Eocene inferiore dei Lessini orientali (Vestenanova – Verona). Lavori Soc. ven. Sci. nat., 40: 47-109.
Beschin C., Busulini A., Tessier G., Zorzin R. (2016) – I crostacei associati a coralli nell’Eocene inferiore dell’area di Bolca (Verona e Vicenza, Italia nordorientale). Mem. Mus. civ. St. nat. Verona – 2. serie. Sezione Scienze della Terra, 9: 189 pp.
Beschin C., Busulini A., Calvagno M., Tessier G., Zorzin R. (2017) – Ypresian Decapod Crustacean faunas from the coral-algal environments in the Eastern Lessini Mountains (Vicenza and Verona territory – NE Italy): a comparative analysis. Bull. Soc. geol. France, 188: 13.
Beschin C., Busulini A., Fornaciari E., Papazzoni C.A., Tessier G. (2018) – La fauna di crostacei associati a coralli dell’Eocene superiore di Campolongo di Val Liona (Monti Berici, Vicenza, Italia nordorientale). Boll. Mus. St. nat. Venezia, 69: 129-215.
Beschin C., Busulini A., Tessier G., Zorzin R. (2019) – La fauna di crostacei dell’Eocene superiore di Parona di Verona (Italia nordorientale): nuovi ritrovamenti. Boll. Mus. St. nat. Venezia, 70: 71-142.
Bittner A. (1875) – Die Brachyuren des Vicentinischen Tertiärgebirges. Denkschr. k. Akad. Wiss. Wien, 34: 63-106.
Bittner A. (1883) – Neue Beiträge zur Kenntniss der Brachyuren-Fauna des Alttertiärs von Vicenza und Verona. Denkschr. k. Akad. Wiss. Wien, 46: 299-316.
Blanco-Rambla J.P., Lemaitre R. (1999) – Neocallichirus raymanningi, a new species of ghost shrimp from the northeastern coast of Venezuela (Crustacea: Decapoda: Callianassidae). Proc. Biol. Soc. Washington, 112(4): 768-777.
Busulini A., Tessier G., Beschin C. (2006) – The genus Phlyctenodes A. Milne Edwards, 1862 (Crustacea: Decapoda: Xanthidae) in the Eocene of Europe. Rev. Mexic. Cie. Geol., 23(3): 350-360.
Busulini A., Beschin C., Tessier G. (2012) – Nuovo contributo alla conoscenza dei crostacei decapodi della marna di Possagno (Eocene superiore – Italia settentrionale). Lavori Soc. ven. Sci. nat., 37: 43-72.
Busulini A., Beschin C., Tessier G. (2019) – Nuovo ritrovamento di Faxegalathea valeccensis Beschin, Busulini & Tessier, 2016 nell’Eocene inferiore di Bolca (Verona – Italia nordorientale). Studi e Ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato”, Montecchio Maggiore (Vicenza), 26: 19-22.
Castro P., NgP.K.L., AhyongS.T. (2004) – Phylogeny and systematics of the Trapeziidae Miers, 1886 (Crustacea: Brachyura ), with the description of a new family. Zootaxa, 643: 1-70.
Ceccon L., De Angeli A. (2013) – Segnalazione di decapodi eocenici infestati da parassiti isopodi (Epicaridea) (Vicenza, Italia settentrionale). Lavori Soc. ven. Sci. nat., 38: 83-92.
Checchia-Rispoli G. (1905) – I Crostacei dell’Eocene dei dintorni di Monreale in Provincia di Palermo. Giorn. Sc. nat. ec. Palermo, 25: 309-325.
Cocco F. (1975) – Nota geologica su Altissimo. In: “Valle del Chiampo”, Antologia: 175 pp.
Cocco F., Boscardin M. (2004) – La geologia del territorio. In: Bertacco M., Lora A., Monchelato M. (eds.), Comune di Altissimo, una comunità e le sue contrade. Le origini e la storia: nuove ricerche. Comune di Altissimo: 222 pp.
Dainelli G. (1915) – L’Eocene friulano. Monografia geologica e paleontologica. Firenze: 721 pp.
De Angeli A., Beschin C. (2008) – Crostacei decapodi dell’Oligocene di Soghe e Valmarana (Monti Berici,Vicenza – Italia settentrionale). Studi e Ricerche – Assoc. Amici Mus. – Mus. civ. “G. Zannato” – Montecchio Maggiore (Vicenza), 15: 15-39.
De Angeli A., Caporiondo F. (2016) – Un nuovo Parthenopidae (Crustacea, Decapoda, Brachyura) dell’Eocene inferiore dei Monti Lessini orientali (Verona – Italia settentrionale). Lavori Soc. ven. Sci. nat., 41: 137-144.
De Angeli A., Caporiondo F. (2017) – I granchi eremiti (Crustacea, Decapoda, Anomura, Paguroidea) dell’Eocene medio di cava “Main” di Arzignano (Vicenza, Italia settentrionale). Studi Trentini Sci. nat., 96: 11-32.
De Angeli A., Caporiondo F. (2020) – Braggitrapezia alessandroi nov. gen., nov. sp. (Crustacea, Decapoda, Trapezioidea) dell’Eocene dei Monti Lessini orientali (Vestenanova, Verona, Italia nordorientale). Lavori Soc. ven. Sc. nat., 45: 121-127.
De Angeli A., Ceccon L. (2012) – Eouroptychus montemagrensis n. gen., n. sp., (Crustacea, Decapoda, Anomura, Chirostylidae) dell’Eocene inferiore (Ypresiano) di Monte Magrè (Vicenza, Italia settentrionale). Lavori Soc. ven. Sci. nat., 37: 19-24.
De Angeli A., Ceccon L. (2013a) – Latheticocarcinus italicus sp. nov. (Decapoda, Brachyura, Homolidae) dell’Eocene inferiore (Ypresiano) di Monte Magrè (Vicenza, Italia settentrionale). Lavori Soc. ven. Sci. nat., 38: 103-109.
De Angeli A., Ceccon L. (2013b) – Tetraliidae and Trapeziidae (Crustacea, Decapoda, Brachyura) from Early Eocene of Monte Magrè (Vicenza, NE Italy). Atti Soc. it. Sc. nat. Mus. civ. St. nat. Milano, 154 (1): 25-40.
De Angeli A., Ceccon L. (2014) – Nuovi brachiuri (Decapoda) dell’Eocene inferiore di Monte Magrè (Vicenza, Italia settentrionale). Lavori Soc. ven. Sci. nat., 39: 77-92.
De Angeli A., Ceccon L. (2015) – Nuovi crostacei brachiuri dell’Eocene di Monte Magrè (Vicenza, Italia settentrionale). Lavori Soc. ven. Sc. nat., 40: 119-138.
De Angeli A., Ceccon L. (2016a) – Montemagralia lata n. gen., n. sp., nuovo crostaceo Trapeziidae (Decapoda, Brachyura) dell’Eocene dei Monti Lessini orientali (Vicenza, Italia settentrionale). Lavori Soc. ven. Sc. nat., 41: 129-136.
De Angeli A., Ceccon L. (2016b) – Paraocalina multilobata Beschin, Busulini, De Angeli & Tessier, 2007 (Crustacea, Brachyura, Xanthidae) nel Terziario del Veneto (Italia settentrionale). Natura vicentina, 19(2015): 5-13.
De Angeli A., Ceccon L. (2017) – Contributo ai crostacei decapodi dell’Eocene inferiore dei Monti Lessini orientali (Italia orientale). Natura vicentina, 20(2016): 5-38.
De Angeli A., Ceccon L. (2020) – Montemagrecarcinus efremi nov. gen., nov. sp., (Decapoda, Brachyura, Pilumnoidea) dell’Eocene inferiore di Monte Magrè (Vicenza, Italia nordorientale). Lavori Soc. ven. Sc. nat., 45: 129-134.
De Angeli A., Garassino A. (2002) – Galatheid, chirostylid and porcellanid decapods (Crustacea, Decapoda, Anomura) from the Eocene and Oligocene of Vicenza (N Italy). Mem. Soc. it. Sci nat. Mus. civ. St. nat. Milano, 30(3): 40 pp.
De Angeli A., Garassino A. (2006) – Catalog and bibliography of the fossil Stomatopoda and Decapoda from Italy. Mem. Soc. it. Sci. nat. Mus. civ. St. nat. Milano, 35 (1): 1-95.
De Angeli A., Garassino A. (2014) – A new genus and species of panopeid crab, Zovocarcinus muelleri (Decapoda, Brachyura), from the Eocene of Zovo di Bolca, Verona (northeast Italy). In: Fraaije R.H.B., Hyžný M., Jagt J.W.M., Krobicki M., Van Bakel B.W.M. (eds.), Proceedings of the 5th Symposium on Mesozoic and Cenozoic Decapod Crustaceans, Krakow, Poland, 2013: A tribute to Pál Mihály Müller. Scripta Geologica, 147: 185-191.
De Angeli A., Garassino A., Pasini G. (2019) – Catalog and bibliography of fossil Stomatopoda and Decapoda from Italy (2007-2018). Mem. Soc. it. Sci. nat. Mus. civ. St. nat. Milano, 45 (1): 1-86.
De Gregorio A. (1895) – Description des faunes tertiaires de la Vénétie. Notes sur certains Crustacés (Brachiures) eocéniques (avec un catalogue de tous les crustacés de la Vénétie cités par les auteurs). Ann. Géol. Pal., 18: 22 pp.
De Zanche V. (1965) – Le microfacies eoceniche nella Valle del Chiampo tra Arzignano e Mussolino (Lessini orientali). Riv. it. Paleont. Stratigr., 71: 925-948.
Di Salvo G. (1933) – I Crostacei del Terziario inferiore della provincia di Palermo. Giorn. Sc. nat. ec. Palermo, 37: 44 pp.
Hyžný M., Karasawa, H. (2012) – How to distinguish Neocallichirus, Sergio, Podocallichirus and Grynaminna (Decapoda: Callianassidae: Callichirinae) from each other in the fossil record ? Bull. Mizunami Fossil Mus., 38: 59-68.
Hyžný M., Zorn I. (2020) – A Catalogue of the Fossil Decapod Crustaceans in the Collections of the Geological Survey of Austria in Vienna. Abh. geol. Bundesanst. Wien, 74, 111 pp.
Karasawa H., Schweitzer C. (2006) – A new classification of the Xanthoidea sensu lato (Crustacea: Decapoda: Brachyura) based on phylogenetic analysis and traditional systematics and evaluation of all fossil Xanthoidea sensu lato. Contr. Zool., 75 (1/2): 23-73.
Kramer M.J., Bellwood D.R., Bellwood O. (2014) – Benthic Crustacea on coral reefs: a quantitative survey. Mar. Ecol. Prog. Ser., 511: 105-116.
Lőrenthey I. (1897) – Adatok Magyarország harmadkorú rákfaunájához. Math. Termész. Ért., 15: 149-169.
Lőrenthey I.(E.) (1898) – Beiträge zur Decapodenfauna des Ungarischen Tertiärs. Termész. Füzetek, 21: 133 pp.
Lőrenthey E., Beurlen K. (1929) – Die fossilen Decapoden der Länder der Ungarischen Krone. Geologica hungarica Ser. Palaeont., 3: 420 pp.
Mietto P. (1997) – Il Paleogene. In: Bagnoli C., Beschin C., Dal Lago A., Mietto P., Piva E., Quaggiotto E. (eds.), Solo a Vicenza – Gli endemismi della provincia. I fossili, le piante, gli animali presenti solo nel territorio vicentino, Blended ed., Vicenza: 173 pp.
Milne EdwardsA. (1861-1865) – Histoire des Crustacés Podophthalmaires fossiles. Paris, Victor Masson et fils, 390 pp.
Milne EdwardsA. (1862) – Monographie des Crustacés fossiles de la famille des Cancériens. Ann. Sci. Nat., Zoologie, s. 4, 18: 31-85.
Müller P., CollinsJ.S.H. (1991) – Late Eocene coral-associated decapods (Crustacea) from Hungary. Contr. Tert. Quatern. Geol., 28 (2-3): 47-92.
NgP.K.L., Guinot D., Davie P.J.F. (2008) – Systema Brachuyrorum: Part I. An annotated checklist of extant Brachyuran crabs of the world. Raffles Bull. Zool., Suppl. 17: 1-286.
Oppenheim P. (1899) – I supposti rapporti dei crostacei terziarii di Ofen descritti da Loerenthey con quelli veneti. Riv. ital. Paleont., 5: 55-62.
Pasini G., Garassino A., De Angeli A., Hyžný M., Giusberti L., Zorzin R. (2019) – Eocene decapod faunas from the Konservat-Lagerstätten laminites of “Pesciara” (Bolca, Verona) and Monte Postale (Altissimo, Vicenza) in northeast Italy: a review and update. N. Jb. Geol. Paläont. Abh., 293(3): 233-270.
Piccoli G. (1966) – Studio geologico del vulcanesimo paleogenico veneto. Mem. Ist. Geol. Min. Univ. Padova, 26: 1-98.
RobinsC.M., Feldmann R.M., Schweitzer C.E. (2013) – Nine new genera and 24 new species of the Munidopsidae (Decapoda: Anomura: Galatheoidea) from the Jurassic Ernstbrunn Limestone of Austria, and notes on fossil munidopsid classification. Ann. Naturhist. Mus. Wien, s. A, 115: 167-251.
Sakai K. (1988) – A new genus and five new species of Callianassidae (Crustacea: Decapoda: Thalassinidea) from Northern Australia. The Beagle, Records of the Northern Territory Museum of Arts and Sciences, 5(1): 51-69.
Schnabel K.E, AhyongT. S. (2010) – A new classification of the Chirostyloidea (Crustacea: Decapoda: Anomura). Zootaxa, 2687: 56-64.
Schweitzer C.E. (2005) – The Trapeziidae and Domeciidae (Decapoda: Brachyura: Xanthoidea) in the fossil record and a new Eocene genus from Baja Califorfnia Sur, Mexico. J. Crust. Biol., 25(4): 625-636.
Schweitzer C.E., Feldmann R.M. (2000) – First notice of the Chirostylidae (Decapoda) in the fossil record and new Tertiary Galatheidae (Decapoda) from the Americas. Bull. Mizunami Fossil Mus., 27: 147-165.
Schweitzer C.E., Feldmann R.M. (2002) – New Eocene decapods (Thalassinidea and Brachyura) from Southern California. J. Crust. Biol., 22: 938-967.
Schweitzer C.E. Feldmann R.M., Gingerich P.D. (2004) – New Decapoda (Crustacea) from the middle and late Eocene of Pakistan and a revision of Lobonotus A. Milne Edwards, 1864. Contr. Mus. Paleont. Univ. Michigan, 31(4): 89-118.
Schweitzer C.E., Gonzáles-Barba G., Feldmann R.M., Waugh D.A. (2005) – Decapoda (Thalassinidea and Paguroidea) from the Eocene Bateque and Tepetate Formation, Baja California Sur, México: systematics, cuticle microstructure, and paleoecology. Ann. Carnegie Mus., 74(4): 275-293.
Schweitzer C.E., Feldmann R.M., Garassino A., Karasawa H., Schweigert G. (2010) – Systematic List of Fossil Decapod Crustacean Species. Crustaceana Monographs, 10: 222 pp.
Schweitzer C.E., Feldmann R.M., Karasawa H. (2012) – Treatise online Number 51 Part R, Revised, Volume 1, Chapter 8M: Systematic Descriptions: Infraorder Brachyura, Section Dromiacea, Paleont. Inst. Kansas Univ.: 43 pp.
Schweitzer C.E., Feldmann R.M., Karasawa H. (2018) – Treatise online Number 112 Part R, Revised, Volume 1, Chapter 8T2: Systematic Descriptions: Superfamily Carpilioidea, Paleont. Inst. Kansas Univ.: 22 pp.
Schweitzer C.E., Feldmann R.M., Karasawa H. (2020) – Treatise online Number 131 Part R, Revised, Volume 1, Chapter 8T9: Systematic Descriptions: Superfamily Parthenopoidea, Paleont. Inst. Kansas Univ.: 11 pp.
Tessier G., Beschin C., Busulini A. (2011) – New evidence of coral-associated crustaceans from the Eocene of the Vicenza Lessini (NE Italy). N. Jb. Geol. Paläont. Abh., 260(2): 211-220.
Visonà D. (1974) – Particolarità tettoniche della zona compresa fra la media valle dell’Agno e l’alta valle del Chiampo. Mem. Soc. Geol. Ital., 13: 23-34.

