Riassunto
Nella Foresta del Cansiglio la vegetazione climax è la faggeta. Nel presente studio è stata presa in considerazione la flora micologica delle associazioni fagetali presenti nel SIC-ZPS Foresta del Cansiglio (Regione Veneto) e si sono evidenziati i legami tra il faggio e le specie ad esso associate con numerosissimi macromiceti simbionti. Dalla ricerca ultratrentennale fatta dagli autori e dai dati bibliografici reperiti, ne è scaturita una tabella con 850 taxa completi di fenologia e sito di raccolta. Questa enorme e inaspettata mole di dati è giustificata dalla varietà di associazioni, di specie vegetali, di condizioni climatiche e pedologiche dell’area indagata. Tra le specie censite numerose sono quelle di elevato valore scientifico in quanto rare per la micoflora italiana e perché presenti in numerose Red list europee.
Abstract
Mycological biodiversity in the beech-woods of the Cansiglio Forest
In the Cansiglio Forest the climax vegetation is the beech forest.
In the present study, the mycological flora of the fagetal associations present in the SIC-ZPS Cansiglio Forest (Veneto Region) is considered, showing how the beech and the associated species are connected with several macromycetes. As the result of a research lasted nearly thirty years, a table containing the sampling date and the collection site of 850 taxa has emerged.
This huge amount of data is justified by the variety of associations and plant species, and by the diversity in the climatic and soil conditions that are present in the area.
The analysis revealed that many of the surveyed species are rare considering the Italian mycoflora.
Furthermore, the presence of those species in numerous European Redlists adds scientific value to our findings.
Introduzione
Questo lavoro, che raccoglie i dati di circa 30 anni, è la realizzazione di un progetto nato dall’ interesse che gli autori hanno per il Cansiglio e in particolare per i suoi funghi. Esso va ad integrare la conoscenza della flora micologica del Cansiglio a seguito della pubblicazione di due lavori precedenti: il primo sui macromiceti associati alle comunità vegetali delle zone umide (Bizio & Borsato, 2016) e il secondo sulla flora micologica delle comunità prative (Borsato & Bizio, 2019). Si è ritenuto inoltre di tenere in considerazione anche raccolte poco distanti dai suoi confini amministrativi in quanto la “Foresta” dal punto di vista ecologico-strutturale non ha limiti così precisi. In Fig. 4 sono indicati i codici, le posizioni e le località dei siti oggetto di rilievo.
Gli autori hanno voluto realizzare la lista floristica delle specie esistenti nella Foresta e riconoscere alla comunità vegetale, la faggeta appunto, il ruolo di entità che accoglie e determina tutti gli altri tipi di comunità, in questo caso quella micologica.
Si potrà notare che numerosi macromiceti ectomicorrizici elencati nel presente lavoro sono legati, oltre che al faggio, anche all’abete rosso e all’abete bianco in quanto queste due specie arboree sono intimamente associate al faggio in tutte e tre le comunità fagetali del Cansiglio, come sarà specificato più avanti. La faggeta va perciò considerata come una associazione di varie specie vegetali e non come un bosco monospecifico che, nel corso dei secoli, è stata ed è tutt’ora oggetto di intensa azione antropica che ha portato a modificazioni, seppur lievi, dell’assetto originario.
Considerate la ricchezza naturale e la preziosità di alcuni suoi elementi, la Foresta del Cansiglio è protetta a livello europeo sia dalla Direttiva Habitat che dalla Direttiva Uccelli, e perciò è diventata “SIC-ZPS IT 3230077, Foresta del Cansiglio-Regione Veneto” facente parte di Rete Natura 2000. Questa porzione di Foresta è stata sito di indagine del presente studio.
Area di studio
La Foresta del Cansiglio è la parte più occidentale delle Prealpi Venete-Friulane e fa parte del massiccio del Cansiglio-Cavallo (Cancian et Al., 1985). Dal punto di vista amministrativo il SIC-ZPS appartiene alla Regione Veneto, alle province di Belluno (comuni di Alpago e Tambre) e Treviso (Fregona e Vittorio Veneto) (Buffa & Lasen, 2010). Dal punto di vista morfologico esso occupa un ampio catino, un polje, risultante dalla fusione di unità carsiche minori pianeggianti: Pian Cansiglio, Pian di Valmenera, Campedei, Pian delle Code, in cui è particolarmente attivo il fenomeno carsico (De Nardi, 1978). L’altitudine media di Pian Cansiglio è di circa 1000 m s.l.m., mentre quella dei rilievi che lo circondano (M. Millifret, M. Croseraz) è di circa 1500 m s.l.m.. Le raccolte sono state estese anche alla faggeta del M. Pizzoc (nella parte compresa nel SIC-ZPS) fino ad una altitudine di circa 1400 m s.l.m..
Per comprendere la vegetazione climax (la faggeta appunto) e l’enorme biodiversità micologica che in essa si sviluppa, è fondamentale la conoscenza del tipo di suolo e del clima presenti nell’area oggetto di studio.
I suoli del Cansiglio si sviluppano sia su calcari marnosi (scaglia rossa e scaglia grigia) che su calcari organogeni di età cretacica. I boschi di faggio sono presenti sui versanti che circondano le zone pianeggianti centrali sopra indicate; su questi versanti si osserva una riduzione dello spessore del suolo e un aumento del pH all’aumentare della pendenza e un aumento della sostanza organica rispetto alle zone prative (Garlato & Borsato, 2016).
Dal punto di vista pedologico, i suoli su cui si sviluppano le faggete del Cansiglio vengono classificati nelle seguenti categorie (Garlato & Borsato, 2016):
– WRB 2006: Cutanic Luvisols (Endosiltic), nei versanti calcareo marnosi con pendenza minore;
– WRB 2006: Haplic Cambisols (Calcaric, Orthosiltic), nei versanti calcareo marnosi più ripidi;
– WRB 2006: Endoleptis Cutanic Luvisols, nei versanti su calcari compatti, con pendenza minore;
– WRB 2006: Epileptic Phaeozems (Calcaric), nei versanti ripidi su calcari compatti.
Per quanto riguarda il clima, dalla rielaborazione dei dati forniti dall’ARPAV (Dipartimento regionale sicurezza del territorio e Ufficio validazione dati e climatologia) si sono ottenuti i seguenti diagrammi (Borsato, 2016):
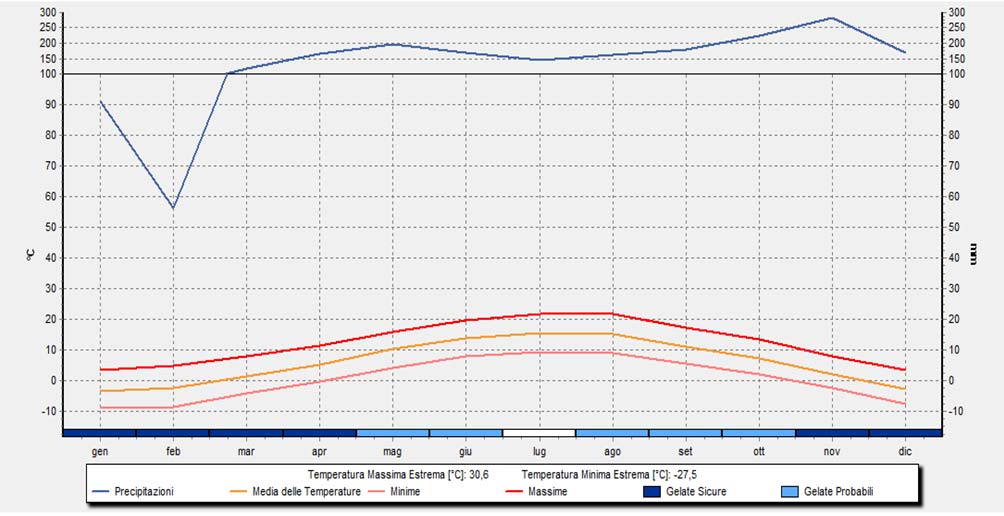
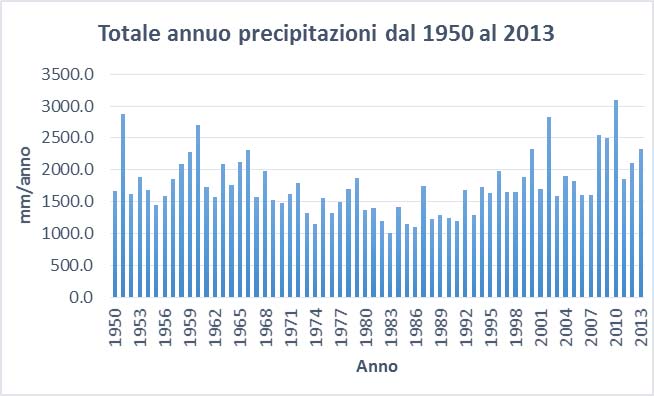
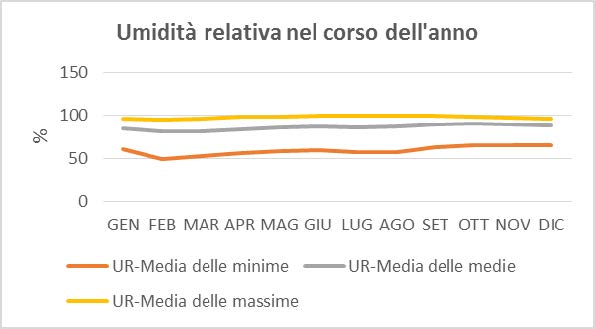
Dall’osservazione dei tre precedenti diagrammi si rileva che:
– Le precipitazioni medie mensili sono superiori a 100 mm/mese quasi tutto l’anno, con un massimo a novembre e un minimo in gennaio-febbraio.
– Le temperature sono tendenzialmente basse con la media del mese più caldo (luglio) di 15.5°C e quella del mese più freddo (gennaio) di -3.7°C.
– La media delle precipitazioni totali annue dal 1950 al 2013 è di 1756.6 mm.
– Durante tutto l’anno l’umidità relativa media oscilla tra l’80-90 %, ne consegue che l’aria del Cansiglio è sempre molto umida e ciò si manifesta con l’abbondanza di precipitazioni nell’area di studio.
Secondo Walter & Lieth (1960), il clima del Cansiglio si definisce umido, con stagione fredda, montano. Esso inoltre è caratterizzato dal fenomeno dell’inversione termica, dovuta alla caratteristica forma a catino dell’altopiano, dentro cui ristagna aria fredda. Ciò viene dimostrato dall’andamento delle gelate che possono avvenire tutto l’anno, tranne che in luglio (Borsato, 2016).
I boschi di faggio del Cansiglio
La faggeta è la comunità vegetale più rappresentativa del Cansiglio, oltre ad essere la sua vegetazione climax. I boschi di faggio sono presenti sui versanti dei rilievi che circondano le zone pianeggianti dell’altipiano, ad una altitudine compresa tra 1000 e 1400 m s.l.m. circa, in contesti freddi e umidi e su pendii generalmente poco inclinati. Sul fondo della conca invece ci sono faggete miste con abete rosso e abete bianco che si accompagnano con boschi puri di peccio di origine antropica (Ubaldi, 2008), i quali spesso sono a contatto con i pratipascoli delle zone pianeggianti o a debole inclinazione, come in Pian Cansiglio.
Da un approfondito studio bibliografico (Poldini & Nardini, 1993; Pignatti E. & Pignatti S., 2014) risulta che le faggete, entro cui sono avvenute le raccolte micologiche studiate nel presente lavoro, si possono attribuire alle associazioni
– Dentario pentaphylli-Fagetum Mayer et Hofmann 1969
– Oxalido- Abietetum Mayer et Hofmann 1969 (Syn.: Abieti-Fagetum Auct.)
– Cardamini trifoliae-Fagetum (Mayer et Hofmann 1969) Oberd. et Müller Th. 1984
come riassunto nel seguente quadro sintassonomico (Sito Internet 5):
71 Cl.: QUERCO ROBORIS-FAGETEA SYLVATICAE Br.-Bl. & Vlieger in Vlieger 1937
71.1 Ord.: FAGETALIA SYLVATICAE Pawłowski in Pawłowski, Sokołowski & Wallisch 1928
71.1.1 All.: Galio odorati-Fagion sylvaticae Knapp ex Tuxen & Oberdorfer 1958 nom. mut.
Oxalido-Abietetum Mayer et Hofmann 1969 (Syn.: Abieti-Fagetum Auct.)
71.1.2 All.: Aremonio agrimonioidis-Fagion sylvaticae (Horvat) Borhidi in Török, Podani & Borhidi 1989
Dentario pentaphylli-Fagetum Mayer et Hofmann 1969
Cardamini trifoliae-Fagetum (Mayer et Hofmann 1969) Oberd. et Müller Th. 1984
Dentario pentaphylli-Fagetum Mayer et Hofmann 1969
La dotazione di specie fagetali illiriche (Cardamine trifolia, Cardamine enneaphyllos,Cyclamen purpurascens) permette, dal punto di vista sintassonomico, di porre questa associazione all’interno dell’alleanza Aremonio agrimonioidis-Fagion sylvaticae dell’ordine Fagetalia sylvaticae (Poldini & Nardini, 1993).
Essa inoltre ha in Cansiglio il locus typicus. Questa foresta ha una struttura a tronchi colonnari alti circa 30 m, è governata a ceduo o a fustaia ed è stabile in condizioni di optimum. Si riproduce facilmente per via gamica soprattutto nelle annate di pasciona (Del Favero R., 2004). Lo strato arboreo è caratterizzato da Fagus sylvatica al quale si associano Abies alba e Picea abies. Durante i mesi estivi i faggi presentano una compatta copertura fogliare che permette una scarsa penetrazione della luce e questo determina la presenza di specie sciafile erbacee e arbustive (Corpo Forestale dello Stato, 2006).
Lo strato arbustivo è composto prevalentemente da giovani individui delle succitate specie arboree, oltre che da Daphne mezereum, Lonicera nigra, Lonicera xylosteum e Rubus idaeus.
Lo strato erbaceo, quando presente, è ricco delle specie tipiche della flora fagetale: Cardamine enneaphyllos, Cardamine bulbifera, Cardamine trifolia, Anemone nemorosa, Luzula nivea, Paris quadrifolia, Prenanthes purpurea, Sanicula europaea. Altre specie nemorali molto diffuse sono Oxalis acetosella e Maianthemum bifolium. Le felci più frequenti sono Athyrium filix-femina, Dryopteris filix-mas, Gymnocarpium dryopteris e Dryopteris carthusiana. Ampi spazi del terreno sono spesso privi di vegetazione erbacea e ricoperti invece da una lettiera costituita da molti strati sovrapposti di foglie di faggio. E’ soprattutto su questo substrato, sul legname in decomposizione, sulle radici affioranti degli alberi e su qualche porzione di terriccio scoperto che vivono i funghi oggetto di questa ricerca.
Questa associazione è presente sulle pendici del M. Millifret e del M. Pizzoc, nella Riserva Naturale biogenetica Campo di Mezzo-Pian Parrocchia, in Crosetta e in Candaglia.
Questa comunità forestale rappresenta un grande valore, sia economico che dal punto di vista del mantenimento dell’equilibrio ambientale e della biodiversità.
Oxalido-Abietetum Mayer et Hofmann 1969 (Syn.: Abieti-Fagetum Auct.)
Il bosco di faggio, nel fondo della conca del Cansiglio, si arricchisce di molti individui di Picea abies e di Abies alba, a causa della presenza di un clima più freddo e umido dovuto al fenomeno dell’inversione termica, con nebbie mattutine e serali. E’ frequente la presenza di rocce affioranti dalla superficie che favoriscono la risalita di aria umida con l’effetto di abbassare ulteriormente la temperatura (Pignatti E. & Pignatti S., 2014). Questa faggeta assume l’aspetto fisionomico dell’Abieti-Fagetum (Poldini & Nardini, 1993), ora noto con il sinonimo Oxalido-Abietetum Mayer et Hofmann 1969.
In questi boschi lo strato arboreo è caratterizzato da Picea abies, Abies alba e Fagus sylvatica, i cui individui spesso non raggiungono l’altezza delle due specie precedenti. Il sottobosco è molto denso, con lo strato erbaceo che comprende specie come Prenanthes purpurea, Petasites albus, Polygonatum verticillatum,Anemone trifolia, Maianthemun bifolium, Oxalis acetosella. Quest’ultima è presente ovunque con alte coperture, assieme ad Anemone nemorosa e Cardamine trifolia. Le felci sono molto abbondanti: Athyrium filix-femina, Dryopteris filix-mas, Gymnocarpium robertianum e Dryopteris carthusiana. Nel complesso la flora del sottobosco è quella delle foreste di faggio, con pochi elementi delle foreste di abete rosso, come le specie di Vaccinium (Pignatti E. & Pignatti S., 2014).
Cardamini trifoliae-Fagetum (Mayer et Hofmann 1969) Oberd. et Müller Th. 1984
In Cansiglio questa associazione si sviluppa sull’altopiano, nelle zone dove il fenomeno carsico è molto evidente e generalmente è esposta verso nord, in ambienti freddi e umidi, caratterizzati da frequenti nebbie. Si presenta come un consorzio di alberi ad alto fusto, spesso con prevalenza di conifere come Abies alba e Picea abies, in cui Fagus sylvatica è presente con coperture minori.
Lo strato arbustivo è ben sviluppato, ma consiste per la massima parte di giovani individui delle specie arboree sopraindicate; inoltre sono presenti Lonicera nigra e Sorbus aucuparia. Lo strato erbaceo è ricco di specie nemorali come Anemone trifolia, Cardamine trifolia, Oxalis acetosella, Petasites albus, Sanicula europaea, Anemone nemorosa e di felci come Athyrium filix-femina, Dryopteris filix-mas e Dryopteris dilatata.
Queste tre diverse comunità vegetali, prese nel loro insieme con la loro varietà di condizioni climatiche, pedologiche, ecologiche e di azione antropica, accolgono la ricchissima flora micologica oggetto del presente studio. Poiché i dati sono stati raccolti nel corso degli ultimi 30 anni (quando gli autori non pensavano a questa pubblicazione) e alcuni altri sono stati desunti dalla bibliografia (per lo più quelli afferenti a Polyporacae s.l. e Corticiaceae s.l.), non ci è stato possibile risalire a quale delle associazioni fagetali appartengano le specie tabellate.
Occorre infine precisare che, per gli stessi dati desunti dalla letteratura, non è stato possibile indicare la fenologia.
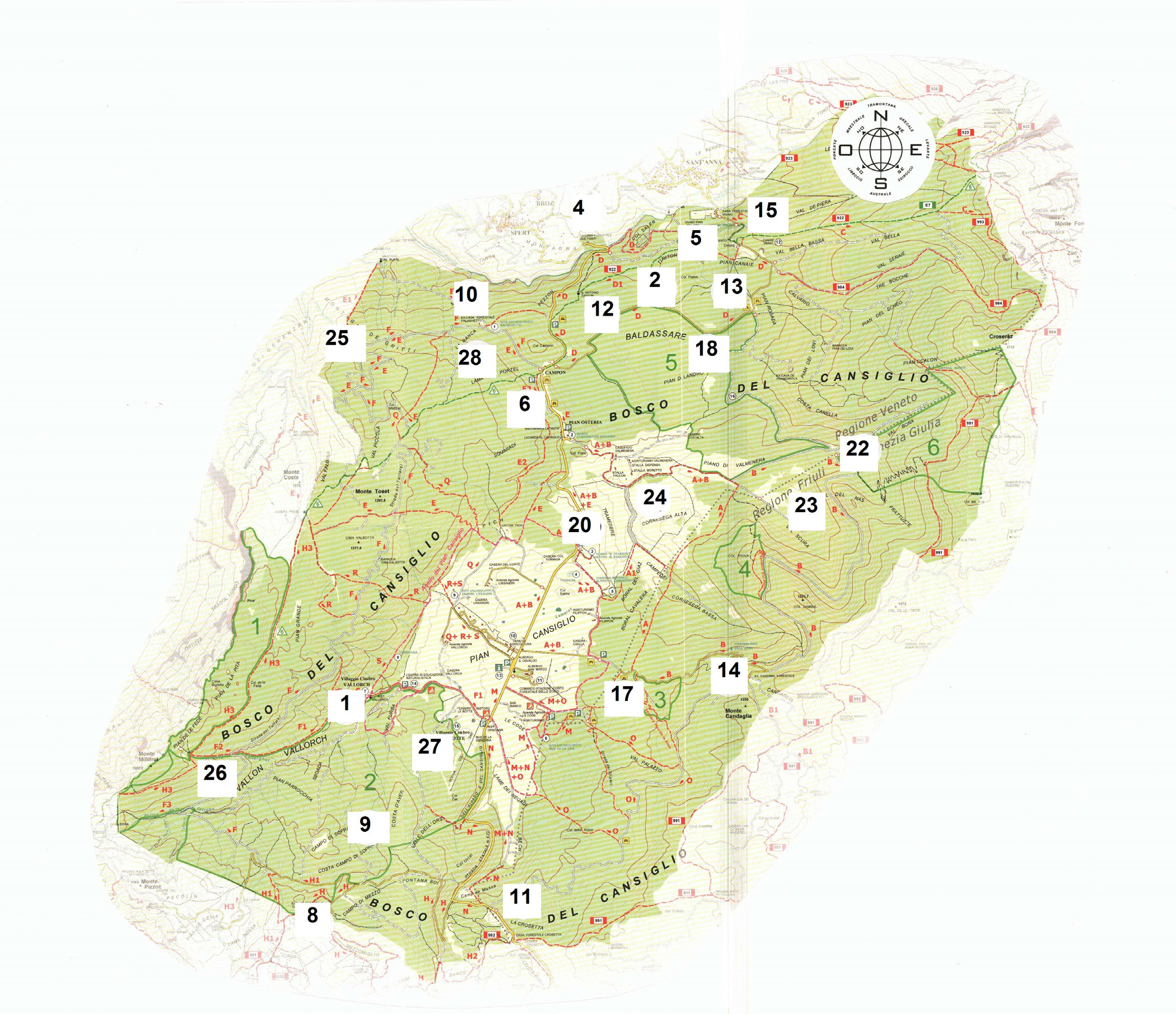
1: Vallorch, 2: Busa della Neve, 4: Due Ponti, 5: Pian dei Spin, 6: Pian Osteria, 8: Cadolten, 9: Costa Campo di Sopra, 10: Palughetto, 11: Crosetta, 12: Val Tritton, 13: Pian Rosada, 14: Candaglia, 15: Vivaio, 17: Strada per Casa Candaglia, 18: Riserva Pian di Landro Baldassare, 20: Tramedere, 22: Valbona, 23: Col Piova, 24: Cornesega alta, 25: M. dei Gritti, 26: Strada Taffarel, 27: Le Rotte, 28: La Banca – Palughetto.
Materiali e metodi
Il progetto di indagine micologica si è sviluppato rilevando i corpi fruttiferi di macromiceti presenti in 23 località rappresentative delle faggete del Cansiglio, più volte indagate negli ultimi 30 anni, fino a tutto il 2018.
In un’ottica di fornire una maggiore compiutezza, l’elenco dei macromiceti pubblicato in questo lavoro si avvale di due contributi che, per la loro rilevanza, abbiamo ritenuto indispensabile acquisire. Il primo contributo è costituito dal censimento eseguito, nelle stesse aree di ricerca e fin dall’inizio degli anni ’90 del secolo scorso, dalla Società Veneziana di Micologia (associazione di cui fanno parte due degli autori del presente lavoro, E.C. ed E.B.). Il secondo è rappresentato dal gruppo dei tre lavori specialistici di Bernicchia e Padovan (1987), di Bernicchia (1988) e di Ubaldi, Bernicchia, Padovan e Zanotti (1990).
La tabella micofloristica, che contiene 850 taxa, è stata concepita in modo da poter offrire un numero ragionevole di dati significativi. In essa vengono riportati, oltre al nome delle specie (in ordine alfabetico) e degli autori, il riferimento numerico della località, la fenologia espressa nei mesi di rilevamento, l’appartenenza ai rispettivi gruppi trofici e tassonomici, il numero delle segnalazioni e la presenza nelle Red list europee prese in considerazione. Si precisa che per le specie tratte dai lavori di Bernicchia, Padovan, Ubaldi e Zanotti (op. cit.) non è stato possibile risalire alla fenologia.
I corpi fruttiferi rilevati sono stati fotografati in ambiente con fotocamere digitali Canon EOS 550D, Fujifilm X20, Nikon D70 e D5300 e identificati prevalentemente da freschi sulla base di osservazioni macro e micromorfologiche, chimiche, organolettiche ed ecologiche. Per i rilievi microscopici sono stati utilizzati microscopi binoculare WILD M11 montante ottiche Leitz-Zeiss e trinoculare Zeiss Primo Star. I preparati sono stati montati in acqua distillata, blu cotone (per evidenziare l’eventuale cianofilia e le ornamentazioni sporali negli ascomiceti), blu di cresile (per la verifica della metacromasia), carminio acetico (per fare risaltare le granulazioni siderofile), floxina (per i tessuti degli ascomiceti), KOH al 5%, reagente di Melzer (per evidenziare l’eventuale amiloidia/destrinoidia), rosso Congo anionico, rosso Congo ammoniacale al 20% (per l’allestimento della maggior parte dei preparati istologici); l’osservazione è avvenuta a 600 e a 1000 ingrandimenti, tramite obiettivo ad immersione.
Alcuni tra i campioni essiccati più significativi sono stati conferiti in MCVE (Erbario Micologico del Museo di Storia Naturale di Venezia) e TUR-A (Erbario Micologico dell’Università di Turku – FIN) e sono a disposizione degli studiosi interessati; altri sono conservati negli erbari personali degli autori. Per la nomenclatura si è fatto riferimento al Codice Internazionale di Nomenclatura (C.I.N.) adottato nel Congresso di Melbourne (2012) (sito internet 1); per l’indicazione del nome corretto, si è utilizzato l’Index Fungorum (sito internet 2), salvo diverso avviso da parte nostra. Le abbreviazioni dei nomi degli autori sono tratte da Kirk & Ansell (2003).
Per il riconoscimento delle specie fungine è stata consultata la letteratura specializzata allo studio dei funghi del piano montano, dell’arco alpino, dell’emisfero boreale, del Nord Europa (Knudsen & Vesterholt, 2008), oltre alle più rilevanti opere di carattere monografico di volta in volta citate nel testo.
Risultati
Le ricerche micologiche hanno avuto come risultato l’osservazione, lo studio e la catalogazione di 3482 segnalazioni in circa 30 anni che hanno portato alla individuazione di 850 diversi taxa fungini. In conformità con la maggior parte delle ricerche finalizzate alla conoscenza della flora micologica di un dato territorio, i risultati si sono incentrati sulla presenza delle diverse specie, trascurando il dato quantitativo.
Appartenenza ai gruppi sistematici: la stragrande maggioranza dei funghi rinvenuti nelle faggete del Cansiglio appartiene al Phylum Basidiomycota (basidiomiceti): a questo gruppo appartengono 751 taxa su 850, pari al 88,4% del totale. Dei rimanenti 87 taxa, pari al 10,2%, appartengono al Phylum Ascomycota (ascomiceti), mentre solo 12, pari all’ 1,4% appartengono al Phylum Myxomycota (mixomiceti, di cui si dirà brevemente in un capitolo a parte).
Appartenenza ai gruppi trofici: in base alle nostre osservazioni e conoscenze dirette, oltre che ai dati di letteratura, è possibile distribuire i funghi delle faggete del Cansiglio nei tre classici gruppi trofici e in ulteriori due gruppi misti, come di seguito:
Saprotrofi (S) 467 taxa, pari al 55% del totale
Ectomicorrizici (E) 341 taxa, pari al 40% del totale
Parassiti (P) 22 taxa, pari al 2,6% del totale
Sapro-ectomicorrizici (S-E) 5 taxa, pari allo 0,6% del totale
Sapro-parassiti (S-P) 3 taxa, pari allo 0,4% del totale
A questi devono essere aggiunti 12 taxa batteriofagi (B), pari al 1,4% del totale, interamente rappresentati dai mixomiceti.
Elenco tassomomico ragionato
– Amanita ochraceomaculata Neville, Poumarat & Fraiture – Questa specie della Sezione Vaginatae si distingue per l’habitat sotto abete rosso, per la tinta nocciola e per il velo di colore giallo-ocraceo che si lacera in scaglie poligonali (Alpago Novello, 2006). A dispetto della sua istituzione recente (Neville et Al. 2000, ne descrivono la raccolta tipica proveniente dalla Val Belluna) si tratta di una specie piuttosto frequente, addirittura comune nell’Agordino, presente in due stazioni in Cansiglio.
– Artomyces pyxidatus (Pers.) Jülich – Abbiamo trovato una sola volta a Vallorch questa inconfondibile “clavaria” caratterizzata per l’aspetto vetroso dei basidiomi e per le ramificazioni terminanti con una piccola coppa coronata. Questo raro saprotrofo lignicolo è inserito in numerose liste rosse dei macromiceti europei.
– Calocybe gangraenosa (Fr.) V. Hofst., Moncalvo, Redhead & Vilgalys – Inserita per lungo tempo nel genere Lyophyllum, è stata di recente ricombinata in Calocybe (2012) in seguito ad indagini genetiche. Si tratta di una specie con scarsissime segnalazioni per il territorio nazionale, caratterizzata da un portamento tricolomatoide, da colorazioni generali biancastre, dal cappello fortemente fibrilloso e dall’annerimento di tutte le parti in seguito a manipolazione o contusioni.
– Calocybe ionides (Bull.) Donk – Conosciuta anche come Rugosomyces ionides (Bull.: Fr.) Bon, è una specie molto rara, trovata una sola volta in Cansiglio e presente in numerose liste rosse europee; legata ai boschi di conifere e misti, è nota per il singolare e sorprendente cromatismo di cui è dotata.
– Caloscypha fulgens (Pers.) Boud. – Genere monospecifico per questa specie capace, con l’excipulum di colore blu-verde e dell’imenio giallo-arancio, di un cromatismo spettacolare. Si tratta di un ascomicete primaverile per il quale è ormai accertata la sua natura di parassita dei semi di Abies alba. Rinvenuto in due stazioni in Cansiglio (faggete con presenza di abete bianco), è considerata specie minacciata in ben undici delle liste rosse europee da noi consultate.
– Cortinarius fervidus P.D. Orton – Inserito nella Sezione Dermocybe per la presenza di particolari pigmenti idrosolubili, C. fervidus si riconosce per la media taglia, il cappello e le lamelle rosso aranciate e per la presenza di un abbondante velo rossastro distribuito a bande sul gambo. È una specie ampiamente distribuita nelle foreste boreali ma estremamente rara nel sud Europa; oltre alla nostra segnalazione, per l’Italia sono noti alcuni sporadici ritrovamenti in Alto Adige.
– Cotylidia pannosa (Sowerby) D.A. Reid – Specie dal portamento piuttosto caratteristico, si presenta sotto forma di rosette vellutate e concresciute di colore fulvo-aranciato, tipicamente su lettiera nei boschi misti di Fagus, Abies e Picea. Nonostante l’aspetto richiami il Genere Thelephora, con questo non ha alcuna affinità, ma geneticamente appartiene a un clade di funghi imenochetoidi in stretta relazione con il genere lamellato Rickenella (Kout & Zìbarova, 2013).
– Dentipellis fragilis (Pers.) Donk – Spettacolare specie caratterizzata da una cascata di aghi che superano spesso la lunghezza di 10 mm, inizialmente di aspetto vetroso, prima biancastri poi ocra-giallastri. Si sviluppa come saprotrofo principalmente su legno di Fagus; nonostante le colonie a volte piuttosto estese, la sua localizzazione non è agevole a causa della crescita sulla parte adiacente al suolo. Considerata la sua presenza in ben 12 Red list europee, si tratta senza dubbio di una specie da proteggere.
– Entocybe turbida (Fr.) T.J. Baroni, V. Hofst. & Largent – Questa specie, tradizionalmente compresa nel genere Entoloma, è stata recentemente (Baroni et Al., 2011) trasferita, assieme ad altre, al nuovo genere Entocybe sulla base di analisi molecolari, sempre all’interno del clade Entomolataceae: il nome del genere, nel combinare Entoloma e Rhodocybe, sottintende le somiglianze con le specie di quei due generi. E. turbida è stata trovata una sola volta a Vallorch, in bosco di faggio misto a peccio.
– Hydnotria tulasnei (Berk.) Berk. & Broome – Curiosa Pezizales globosa, ipogea, affiorante sul terreno di boschi umidi di conifere e di latifoglie. Rinvenuta un paio di volte in località Busa della Neve, si riconosce piuttosto facilmente alla sezione per la sua gleba meandriforme, lacunosa, cioè con numerose cellette sinuose separate da venature ocra-rossastre.
– Hygrophorus hyacinthinus Quél. – Per portamento e colorazioni, potremo definire H. hyacinthjnus una specie anonima, ciò che la rende inconfondibile è il profumo dolcissimo e floreale (di giacinto come suggerisce il nome) che emana l’esemplare giovane; gli esemplari troppo maturi modificano il loro odore diventando sgradevolmente terroso. È una specie poco diffusa a distribuzione prevalentemente centro europea, molto rara al nord; ciò motiva la sua presenza in numerose Red list europee.
– Infundibulicybe mediterranea Vizzini, Contu & Musumeci – Ciò che rende interessanti e sorprendenti le nostre segnalazioni è l’ambiente di crescita che per questa specie è tipicamente mediterraneo. Separata inizialmente a livello varietale dalla “capogruppo” Infundibulicybe gibba (Pers.) Harmaja, ha assunto la dignità specifica in seguito ad indagini biomolecolari (Vizzini & Al., 2011). Oltre che per aspetti genetici, la validità della specie è sostenuta dalla costanza delle colorazioni bruno-rossastre, dalle dimensioni sporali inferiori e per la reazione verdastra all’idrossido di potassio.
– Inocybe sphagnophila Bandini & B. Oertel – L’unica raccolta del Cansiglio (Campo di Sopra) è stata classificata grazie alla perfetta coincidenza degli elementi morfologici ed ecologici con questa specie della sez. Napipedinae istituita di recente dalla Germania (Bandini et Al., 2017), dove è stata trovata più volte in luoghi umidi con presenza di sfagni, sempre in compagnia di abeti rossi o faggi. Un precedente ritrovamento (E.B.) proviene dalla Foresta di Paneveggio, sotto abeti rossi, ma sempre con sfagni.
– Isaria farinosa (Holmsk.) Fr. – Il genere Isaria comprende ascomiceti entomopatogeni, cioè parassiti di insetti, e rappresenta l’anamorfo (stadio riproduttivo asessuato, spesso simile ad una muffa), il cui teleomorfo (stadio riproduttivo sessuale) risulta appartenere al genere Ophiocordyceps. In particolare Isaria farinosa è un agente patogeno di larve e adulti di parecchi insetti, tra cui il genere di lepidotteri Thitarodes degli altopiani tibetano e himalayano che ospitano il teleomorfo Ophiocordyceps sinensis, specie classificata tra i funghi medicinali cinesi col nome comune di “fungo bruco” e tanto sfruttata da dover essere la sua raccolta regolamentata dalla legge cinese. Le nostre raccolte di I. farinosa, sono stati reperiti su adulto di vespa e di carabide non identificato.
– Kuehneromyces lignicola (Peck) Redhead – Si tratta di una specie precoce macroscopicamente separabile con difficoltà da Galerina marginata s.l.; con quest’ultima condivide la crescita su ceppaie di conifera, l’anello piccolo e spesso fugace, nonché la fibrillosità del gambo. Ha una distribuzione prevalentemente nord europea e la sua presenza in Cansiglio può essere imputata al fatto che le condizioni climatiche simulano per gran parte dell’anno quelle boreali. Rare le segnalazioni per il territorio nazionale.
– Lepiota fuscovinacea F.H. Møller & J.E. Lange – Può essere senz’altro considerata una specie rara, ben caratterizzata sotto il profilo microscopico per l’unicità dei caratteri ma identificabile anche macroscopicamente per le colorazioni generali grigio-violacee, la decorazione del cappello tomentoso-lanosa e non chiaramente squamata e per il gambo concolore e cotonoso. Cresce su lettiera di numerose conifere, dai boschi subalpini ai parchi cittadini.
– Leucangium carthusianum (Tul. & C. Tul.) Paol. – Più conosciuto come Picoa carthusiana Tul. & C. Tul., è un ascomicete ipogeo, spesso affiorante sul terreno nei boschi misti. Raccolto una sola volta in Val Tritton nel mese di novembre, questa specie che assomiglia ad un tartufo del genere Tuber, è probabilmente meno rara di quanto i suoi ritrovamenti facciano presumere.
– Ophiocordyceps ditmarii (Quél.) G.H. Sung, J.M. Sung, Hywel-Jones & Spatafora – Come già chiarito in precedenza (vedi I. farinosa), il genere Ophiocordyceps annovera specie prevalentemente entomopatogene, che possono aggredire sia larve che adulti. La specie in oggetto era nota nel passato come O.sphaecocephala (Klotzsch ex Berk.) Berk. & M.A. Curtis 1868, ma è ora appurato che questa specie (morfologicamente molto simile) ha una distribuzione esclusivamente tropicale, a differenza di O. ditmarii che è invece distribuita in Europa (Van Vooren & Audibert, 2005). Sia gli esemplari dei due ritrovamenti del Cansiglio che altri quattro effettuati nei boschi di Palù S. Marco nel comune di Auronzo (E.C.), si sviluppavano su vespe adulte.
– Peziza saniosa Schrad. – Si tratta di una peziza poco frequente nei boschi di faggio (predilige le conifere), trovata solo due volte in Cansiglio e presente in sei delle liste rosse europee da noi consultate. Nonostante la rarità, non è difficile determinare questa specie perché produce ascomi di colore blu-nero, che alla frattura emettono un lattice blu-viola brillante.
– Podostroma leucopus P. Karst. – Ridefinito recentemente il genere, oggi Podostroma annovera al suo interno alcune specie stipitate del genere Hypocrea; l’unica specie europea con cui potrebbe essere confuso è P. alutaceum distinguibile principalmente per la crescita lignicola e non terricola. Tende inoltre a sviluppare ascomi a forma di clava regolare anziché presentare una netta strozzatura tra parte fertile e sterile come invece accade in P. leucopus.
– Psathyrella caput-medusae (Fr.) Konrad & Maubl. – Specie rinvenuta, come anche numerose altre commentate in questo elenco, nella cosiddetta “Busa della Neve”, località evidentemente molto favorevole alla crescita di tante specie interessanti. P. caput-medusae cresce a ciuffi, in autunno, su resti legnosi di latifoglie e di conifere; si riconosce agevolmente per le squame irte di colore bruno “caldo”, per l’anello persistente sul gambo, per l’odore aromatico, dolciastro.
– Russula anthracina var. insipida Romagn. – Entità appartenente alla omogenea Sezione Compactae Fr. (specie provvieste di lamellule e fortemente imbrunenti/annerenti con l’età o alla manipolazione) R. anthracina si riconosce per il cappello subvellutato, le lamelle estremamente fitte con evidenti tonalità rosate, l’annerimento delle parti senza un preventivo arrossamento e sapore pepato.
Per quanto riguarda la var. insipida si differenzia per il sapore dolce e le spore più reticolate.
– Russula cuprea Krombh. – Quest’unico ritrovamento ci ha lasciati un po’ perplessi, abituati a reperire questa specie nei querceti caldi collinari, ma dalla letteratura è stato appurato che può instaurare legami micorrizici anche con il faggio (Sarnari, 1998). È un’entità della Subsezione Urentes Maire, che raggruppa specie pepate a sporata gialla; trattandosi di un raggruppamento eterogeneo si è resa necessaria un’ulteriore suddivisione in Serie di cui R. cuprea è la capofila (Serie Cuprea). La distinzione dalle altre 5 specie è affidata principalmente alle dimensioni sporali (le più piccole della Serie) e all’estrema variabilità cromatica del cappello.
– Russula faustiana Sarnari – Specie strettamente legata al faggio, di colore grigio o grigio-nocciola con sapore mite, ascrivibile alla Subsezione Griseinae J. Schaeffer. Dalle consimili si separa in primis per l’assenza di tonalità verdi sul cappello. Al pari di altre specie (es. Amanita virosa, Strobilomyces strobilaceus) sembra sia in espansione nella Foresta del Cansiglio, probabilmente dovuta all’età della foresta che determina un diverso microclima del sottobosco ed una variazione del pH del suolo. Durante la nostra indagine è stata ritrovata quattro volte in località Busa della Neve.
– Strobilomyces strobilaceus (Scop.) Berk. – Questo boleto inconfondibile per le squame fioccose grigio-nerastre era da considerarsi raro o poco frequente, tanto da risultare inserito in ben nove liste rosse dei macromiceti europei. Tuttavia noi l’abbiamo rinvenuto più volte in quattro stazioni del Cansiglio, con una frequenza sempre crescente.
– Syzigospora tumefaciens (Ginns & Sunhede) Ginns – Fungo dall’incerta interpretazione sistematica, attualmente sembra accertata l’appartenenza all’ordine delle Tremellales. Produce corpi fruttiferi apparentemente di forma tremelloide, cerebriformi, di color ocra-bruno chiaro sui basidiomi di alcune specie collibioidi (specialmente su Gymnopus dryophilus, ma anche su Gymnopus hariolorum, Gymnopus aquosus e Rhodocollybia butyracea). In realtà Syzigospora è un parassita che, in condizioni di eccessiva umidità, induce l’ospite a produrre delle piccole galle tremelloidi sulle quali produce una sottile pellicola gelatinosa superficiale (Bizio e Losi, 1989). Da noi trovato una volta in località Busa della Neve.
– Tricholoma luridum (Schaeff.) P. Kumm. – Bellissima specie appartenente al gruppo di tricolomi con colori giallo-verdastri; lo caratterizzano la taglia robusta, il cappello con grosse fibrille innate con riflessi metallici e lamelle grigie già nei giovanissimi esemplari. Per ora sono note due sole stazioni a qualche decina di metri una dall’altra, entrambe in località Busa della Neve.
Presenza dei myxomycetes nel censimento micofloristico
Com’è noto, i mixomiceti sono degli esseri viventi del tutto particolari, che si spostano sul substrato come gli animali e si riproducono tramite spore al pari dei funghi. Tuttavia essi non sono né funghi, né animali e, secondo gli studi più recenti, sono considerati un clade a parte negli Unikonta assieme ad animali e funghi. Sin dai tempi più remoti, quando gli aspetti biologici, fisiologici, ecologici e sistematici non erano ancora ben chiariti, i mixomiceti sono sempre stati considerati dei veri e propri funghi, benché mucillaginosi, e come tali sono stati descritti e illustrati in libri e atlanti; questa prassi perdura anche in lavori più recenti.
Nonostante la consapevolezza che tutto ciò non è più sostenibile, in virtù di una scelta arbitraria che avvantaggia l’aspetto documentativo della ricerca, abbiamo optato per il mantenimento, nell’elenco micofloristico, delle 12 specie di mixomiceti rinvenute nelle località di ricerca.
Tab. 1. Tabella micofloristica degli 850 taxa censiti nel presente lavoro.
Legenda gruppi trofici:
S= saprotrofi,
E= ectomicorrizici,
P= parassiti,
S-E= sapro-ectomicorrizici,
S-P= sapro-parassiti,
B= batteriofagi
Legenda gruppi sistematici:
A= Ascomycota,
B= Basidiomycota,
M= Myxomycota
Legenda Red list:
AL= Albania,
AM= Armenia,
BAV= Baviera,
BG= Bulgaria,
CH= Svizzera,
CZ= Rep. Ceca,
EST= Estonia,
FIN= Finlandia,
GB= Gran Bretagna,
HR= Croazia,
IP= Italia Provv.,
IS=Italia ISPRA,
LV= Lettonia,
MNE= Montenegro,
N= Norvegia,
NMK= Macedonia del Nord,
P= Polonia,
RO= Romania,
RUS= Russia,
S= Svezia,
SK= Slovacchia,
SLO= Slovenia.
Conclusioni
L’elenco micofloristico, comprendente 850 taxa rinvenuti all’interno delle faggete del Cansiglio, sebbene con relazioni diverse con Fagus sylvatica, si presta a numerose considerazioni.
Red list. Lo strumento delle liste rosse è stato introdotto dall’Unione Internazionale per la Conservazione della Natura (IUCN) (sito internet 3) che si occupa della conservazione della biodiversità, fornendo anche valutazioni sul rischio di estinzione a livello di specie. Nel corso degli anni e con l’aumento delle conoscenze scientifiche, il ruolo dei funghi come regolatori naturali degli ecosistemi è stato ampiamente riconosciuto e viene prestata maggiore attenzione alla conservazione della micoflora (Siniscalco et al., 2019). Pertanto abbiamo ritenuto utile consultare una ventina di Red list europee di macromiceti ricavabili dall’ European Council for the Conservation of Fungi (sito internet 4). Riportiamo di seguito la legenda delle sigle utilizzate nella tabella: AL= Albania, AM= Armenia, BAV= Baviera, BG= Bulgaria, CH= Svizzera, CZ= Rep. Ceca, EST= Estonia, FIN= Finlandia, GB= Gran Bretagna, HR= Croazia, LV= Lettonia, MNE= Montenegro, N= Norvegia, NMK= Macedonia del Nord, P= Polonia, RO= Romania, RUS= Russia, S= Svezia, SK= Slovacchia, SLO= Slovenia.
Per l’Italia abbiamo consultato la lista rossa provvisoria coordinata da C. Perini (IP) a nome del gruppo di lavoro per la micologia della Società Botanica Italiana (SBI) e la lista rossa (IS) prodotta dall’ISPRA (Siniscalco et al., 2019).
Ambedue sono da considerarsi largamente provvisorie e tutt’altro che esaustive; la seconda è, come riconosciuto dagli stessi Autori, una sorta di “work in progress.”
Successivamente abbiamo confrontato le specie rinvenute nelle faggete del Cansiglio con tali liste rosse europee e il risultato è stato che ben 428 specie su 850 (dunque oltre la metà) è presente in una o più delle sopra citate Red list.
Questo semplice dato conferisce, da solo, un’oggettiva rilevanza alla nostra indagine: il numero dei macromiceti e il loro “peso specifico” in termini ecologici, confermano che le faggete del Cansiglio costituiscono un ambiente di grande pregio non solo dal punto di vista paesaggistico e naturalistico in senso lato, ma anche, e soprattutto, sotto l’aspetto della biodiversità.
In particolare, si può notare che circa quindici specie comprese nel censimento con un modesto numero di campionamenti, si confermano rare o a rischio anche in altri paesi europei, tanto da essere presenti contemporaneamente in un elevato numero di Red list. È forse più interessante constatare che parecchie specie, non rare o addirittura frequenti nelle faggete del Cansiglio, sono invece considerate rare, poco frequenti o minacciate in altri paesi europei.
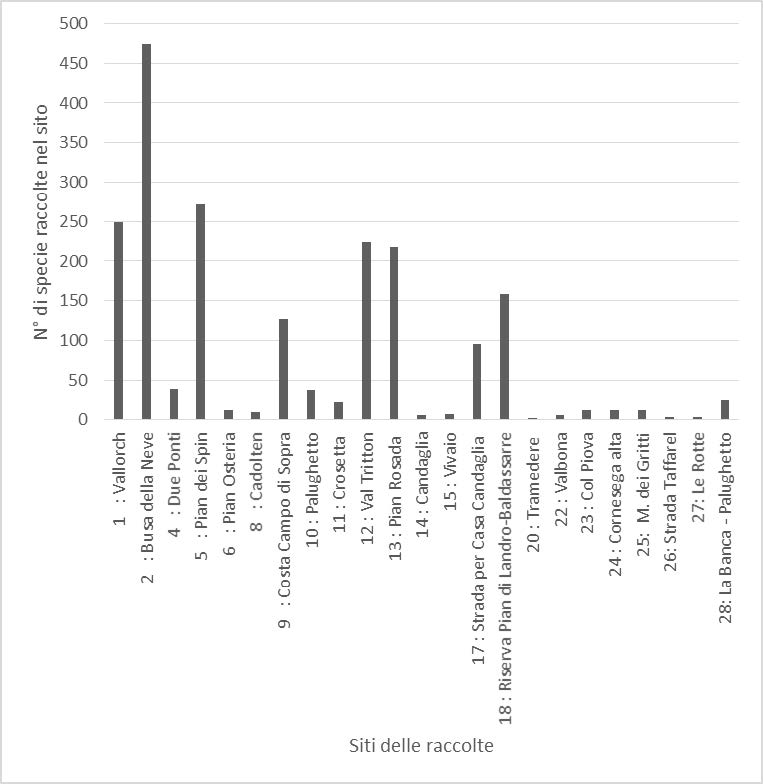
Concludendo, per quanto riguarda la ricchezza micologica dei singoli siti (calcolata in modo quantitativo come numero di specie per sito), si può notare dal seguente grafico come la Busa della Neve detenga il primato con 474 taxa presenti; ad essa seguono Pian dei Spin (272), Vallorch (249), Val Tritton (224) e Pian Rosada (218). Tutte queste località sono caratterizzate anche da una notevole biodiversità floristica, in cui al faggio si accompagnano l’abete rosso e l’abete bianco, oltre che un notevole numero di specie erbacee e arbustive.
Ringraziamenti
Si ringraziano i Sigg. A. Bernicchia, F. Padovan, D. Uboldi e A. L. Zanotti, autori di tre fondamentali contributi sui funghi lignicoli delle faggete del Cansiglio dai quali abbiamo acquisito dati e informazioni utili alla stesura del nostro lavoro.
Per le ricerche sul campo si ringrazia la Società Veneziana di Micologia nelle persone dei seguenti micologi che ne hanno fatto parte durante la sua trentennale attività: D. Alzani, F. Bersan, A. Bizzi, A. Camoli, M. Castoldi, E. Cautero (†), R. Cerello, F. Ferigo, L. Levorato, C. Losi, M. Marchetti, G. F. Medardi, G. Pegoraro, P. Popolizio (†), G. Ricci (†), G. Robich, R. Tassan Toffola, L. Zambolin, G. Zecchin; inoltre i Sigg. G. Altinier, C. Angelini, V. Antoniolli, I. Barbiero, A. Coan (†), S. Florian, C. Gallinaro, M.C. Magnozzi, R. Pavan, D. Signorini.
Si ringraziano i Sigg. H. e R. Grünert per il contributo alla conoscenza della Red list della Baviera.
Un sentito ringraziamento a Raffaella Trabucco e Jukka Vauras, rispettivamente curatori degli erbari MCVE e TUR-A.
Un grazie riconoscente al Prof. Em. Livio Poldini e al Prof. Michele Codogno dell’Università di Trieste.
Bibliografia citata
Alpago Novello L. (2006) – Funghi rari o poco noti della Sinistra Piave in Valbelluna. Tipografia Milani, Verona.
Arpav, Dipartimento Regionale sicurezza del Territorio, Servizio Meteorologico: dati meteorologici 1950-2013.
Arpav, Servizio meteorologico, Ufficio Validazione dati e Climatologia (2013) – Le principali variabili meteorologiche del Cansiglio.
Bandini D., Christan J., Eberhardt U., Ploch S., Tahir A., Oertel B., Thines M. (2017) – Inocybe sphagnophila sp. nov., a new species of the nodulose-spored subsection Napipedinae of genus Inocybe (Agaricales). Mycol. Bav. 18: 11-34.
Baroni T., Hofstetter V., Largent D., Vilgalys R. (2011) – Entocybe is proposed as a new genus in the Entolomataceae (Agaricomycetes, Basidiomycota) based on morphological and molecular evidence. North American Fungi. 6: 1-19.
Bernicchia A. (1988) – Aphyllophorales del Bosco del Cansiglio: Polyporaceae s.l. Mic. e Veg. Medit. 3(1): 11-22.
Bernicchia A., Padovan F. (1987) – Aphyllophorales del Bosco del Cansiglio: Corticiaceae s.l. Mic. e Veg. Medit. Vol. II(1): 25-36.
Bizio E., Borsato V. (2016) – Indagine preliminare dei macromiceti associati alle comunità vegetali delle zone umide della Foresta del Cansiglio (Veneto-NE-Italia). Lavori Soc. Ven. Sc. Nat. 41: 87-113.
Bizio E., Losi C. (1989) – Due rari Basidiomycetes della Valle del Biois (BL) Syzygospora tumefaciens (Ginns & Sunhede) Ginns e Femsjonia peziziformis (Lév.) Karst. Riv. di Micol. 3-4: 181-189.
Borsato V. (2016) – Il SIC-ZPS Foresta del Cansiglio e la sua biodiversità. Pubblicazione in proprio.
Borsato V., Bizio E. (2019) – La flora micologica delle comunità prative della Foresta del Cansiglio (Veneto, NE Italy). Lavori Soc. Ven. Sc. Nat. 44: 49-66.
Buffa G., Lasen C. (2010) – Atlante dei siti Natura 2000 del Veneto. Regione del Veneto-Direzione Pianificazione Territoriale e Parchi, Venezia, pp. 394.
Cancian G., Ghetti S., Semenza E. (1985) – Aspetti geologici dell’Altopiano del Cansiglio, Lavori – Società Veneziana di Scienze Naturali – Suppl. 10: 79-90, Venezia, 15-3-1985.
Corpo Forestale dello Stato (2006), Quaderni naturalistici, Archivio del Cansiglio, “Il sottobosco delle Faggete del Cansiglio”, Eurooffset, Maerne di Martellago (VE).
De Nardi A. (1978) – Il Cansiglio Cavallo, lineamenti geologici e morfologici, Doretti UD.
Del Favero R. (2004) – Biodiversità e indicatori nei tipi forestali del Veneto, Multigraf, Spinea (VE). Direttiva n° 79/409/CEE e successiva versione n°2009/147/CEE (Direttiva uccelli).
Direttiva n°92/43/CEE (Direttiva habitat).
Garlato A., Borsato V. (2016) – I suoli del SIC-ZPS IT3230077 “Foresta del Cansiglio” (NE-Italia). Lavori Soc. Ven. Sc. Nat. 41: 115-119.
Kirk P.M. & Ansell A.E. (2003) – Authors of fungal names. A list of authors of scientific names of fungi, with recommended standard forms of their names, including abbreviations index of fungi supplement. England, Wallingford: C.A.B. International.
Kout J., Zìbarovà L. (2013) – Revision of the genus Cotylidia (Basidiomycota, Hymenochaetales) in the Czech Republic. Czech Mycology 65(1): 1-13.
Pignatti E., Pignatti S. (2014) – Plant life of the Dolomites. Vegetation structure and ecology, Springer.
Poldini L., Nardini S. (1993) – Boschi di forra, faggete e abieteti in Friuli (NE Italia), Studia geobotanica 13, 215-298.
Sarnari M. (1998) – Monografia illustrata del genere Russula in Europa. Tomo primo. AMB, Centro Studi Micologici, Trento.
Siniscalco C., Lavorato C., Bianco P.M., Rotella M., De Marco V., Floccia F., Campana L. (2019) – Contribution of Ispra “Centers of Excellence” to the development of the Italian Fungal Red List.
Ubaldi D. (2008) – La vegetazione boschiva d’Italia, Cleup.
Ubaldi D., Bernicchia A., Padovan F., Zanotti A.l. (1990) – Osservazioni micofitosociologiche nelle foreste del Cansiglio. Mic. e Veg. Medit. 5(1-2): 25-38.
Van Vooren N., Audibert C. (2005) – Révision du complexe “Cordyceps sphaecocephala” 1re partie: les guepes végétales. Bull. mens. Soc. linn. Lyon 74(7-8): 221-254.
Vizzini A., Contu M., Musumeci E., Ercole E. (2011) – A new taxon in Infundibulicybe gibba complex (Basidiomycota, Agaricales, Tricholomataceae) from Sardinia (Italy). Mycologia 103(1): 203-208.
Walter H., Leith H. (1960) – Klimadiagramm weltatlas, Veb Gustav Fischer Verlag Jena.
Bibliografia consultata
Basso M.T. (1999) – Lactarius Pers. – Fungi Europaei 7. Mykoflora, Alassio.
Boccardo F., Traverso M., Vizzini A., Zotti M. (2008) – Funghi d’Italia. Zanichelli Editore, Bologna.
Bon M. (1990) – Flore Mycologique d’Europe. Les Hygrophores. Documents Mycologiques Mémoire Hors Série n° 1. Amiens.
Bon M. (1992) – Clé monographique des espèces galero-naucorioides. Doc. Mycol., XXI(84): 1-84.
Bon M. (1993) – Flore Mycologique d’Europe. Les Lepiotes. Documents Mycologiques Mémoire hors série n° 3. Amiens.
Bon M. (1997) – Flore Mycologique d’Europe. Clitocybes, Omphales et ressemblants. Documents Mycologiques Mémoire hors série n° 4. Amiens.
Bon M. (1999) – Flore Mycologique d’Europe. Les Collybio-marasmïoïdes et ressemblants. Documents Mycologiques Mémoire hors série n° 5. Amiens.
Breitenbach J., Kränzlin F. (1986) – Champignons de Suisse – vol. 2. Mykologia, Luzern.
Campo E. (2015) – Hygrophorus, Hygrocybe e Cuphophyllus del Friuli Venezia Giulia. Grafiche Scarpis, Sacile.
De Haan A., Walleyn R. (2002) – Studies in Galerina. Galerina Flandriae (1). Fungi non Delineati 23: 1-66.Edizioni Candusso.
De Haan A., Walleyn R. (2006) – Studies in Galerina. Galerina Flandriae (2). Fungi non Delineati 33: 1-73. Edizioni Candusso.
Eyssartier G., Roux P. (2011) – Le guide des Campignons, France et Europe. Ed. Belin, Paris Cedex.
Hausknecht A. (2009) – Conocybe & Pholiotina – Fungi Europaei 11. Edizioni Candusso, Lomazzo.
Julich W. (1989) – Guida alla Determinazione dei Funghi. Vol. 2. Aphyllophorales, Heterobasidiomycetes, Gasteromycetes. Saturnia, Trento.
Knudsen H., Vesterholt J. (2008) – Funga Nordica. Nordsvamp, Copenhagen.
Medardi G. (2006) – Atlante fotografico degli Ascomiceti d’Italia. A.M.B. Fondazione Centro Studi Micologici, Trento.
Ministero dell’Ambiente (2010) – Manuale italiano di interpretazione degli habitat (Direttiva 92/43/CEE).
Montecchi A., Lazzari G. (1993) – Atlante fotografico di Funghi ipogei. A.M.B. Fondazione Centro Studi Micologici, Trento.
Neville P., Poumarat S., Fraiture A. (2000) – Una nuova specie europea di Amanita, Sezione Vaginatae: A. ochraceomaculata. Boll. Gr. Micol. G. Bres. 43(2): 261-268.
Noordeloos M.E. (1992) – Entoloma s.l. – Fungi Europaei 5. Libreria Editrice Giovanna Biella, Saronno.
Noordeloos M.E. (2004) – Entoloma s.l. – Fungi Europaei 5A. Edizioni Candusso, Alassio.
Noordeloos M.E. (2011) – Strophariaceae s.l. – Fungi Europaei 13. Edizioni Candusso, Alassio.
Parra Sànchez L.A. (2008) – Agaricus L. – Fungi Europaei 1. Edizioni Candusso, Lomazzo.
Parra Sànchez L.A. (2013) – Agaricus L. – Fungi Europaei 1A. Candusso Edizioni, Lomazzo.
Robich G. (2003) – Mycena d’Europa – vol.1. A.M.B. Fondazione Centro Studi Micologici, Trento.
Robich G. (2016) – Mycena d’Europa – vol.2. A.M.B. Fondazione Centro Studi Micologici, Trento.
Sarasini M. (2005) – Gasteromiceti epigei. A.M.B. Fondazione Centro Studi Micologici, Trento.
Sarnari M. (2005) – Monografia illustrata del genere Russula in Europa. Tomo secondo. AMB, Centro Studi Micologici, Trento.
Sitografia
1. https://www.iapt-taxon.org/nomen/main.php
2. http://www.indexfungorum.org/
3. https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species
4. http://www.eccf.eu/redlists-en.ehtml
5. www.prodromo-vegetazione-italia.org
6. https://www.nature.com/articles/nmicrobiol201648#s1L aura A. Hug, Brett J. Baker, Karthik Anantharaman, Christopher T. Brown, Alexander J. Probst1, Cindy J. Castelle, Cristina N. Butterfield, Alex W. Hernsdorf, Yuki Amano, KotaroIse, Yohey Suzuki, Natasha Dudek, David A. Relman, Kari M. Finstad, Ronald Amundson, Brian C. Thomas and Jillian F. Banfield (2016) A new view of the tree of life. Nature Microbiology volume 1, Article number: 16048 (2016)
Cartografia
IGM, Edizione 5-1963, Carta topografica d’Italia, scala 1:25000, F.23 II SE Bosco del Cansiglio.
Carta escursionistica 1:25.000 (Centro Caseario Cansiglio, Corpo Forestale dello Stato, Regione veneto).
Carta escursionistica 1:30.000 [(2015, Progetto by ADVERTIME, Imprimenda (PD)].

